Search
Deciphering Nuclease Digestion Data
The properties of an RNA molecule are determined not only by the RNA sequence but also by its final folded structure; structural analysis is integral to understanding the biochemical role of an RNA molecule. To facilitate RNA structural studies, Ambion now offers 3 RNA-grade ribonucleases: RNase A, RNase V1 and RNase T1. These nucleases are ideal for RNA structure determination, sequencing, and protein footprinting or boundary analysis. This article serves as a brief introduction, and includes a hypothetical example, on using nuclease digestion data to deduce the secondary structure of an RNA molecule.
One of the first questions asked about an RNA molecule's secondary structure is, "Which regions are double-stranded?" A good approach to answer this question is to use RNase V1, which preferentially cleaves between nucleotides in double-stranded regions of the RNA. In such an experiment, RNA is end labeled with 32P (e.g., with T4 polynucleotide kinase and [gamma-32P]ATP to label the 5' end or with RNA ligase and [alpha-32P]ATP to label the 3' end), partially digested with RNase V1, and then the fragments are resolved on a denaturing polyacrylamide gel (1-4). For convenience, samples digested with several different enzymes are often analyzed on the same gel. To help identify the cleavage site locations, a "ladder" generated by alkaline hydrolysis or enzymatic digestion is run on the same gel.

To identify or confirm unpaired RNA regions (e.g., loop structures), nucleases selective for single-stranded RNA regions are useful. RNase A, which cleaves 3' to single-stranded C and U residues, and RNase T1, which cleaves single-stranded G residues, are good choices for this type of experiment.
A Hypothetical Example
To choose our hypothetical sample RNA, we analyzed several random RNA sequences with the mfold program developed by Dr. Michael Zuker (5). This program predicts RNA secondary structure based on a complex set of thermodynamic rules. A 54 nt sequence predicted to have a stable secondary structure (-27 kcal at 37°C) was chosen as our "test" RNA.
If the "test" RNA were 5' radiolabeled and digested individually with RNases A, T1 and V1, and the fragments were then analyzed by denaturing gel electrophoresis, the results would resemble the hypothetical data shown in Figure 1.
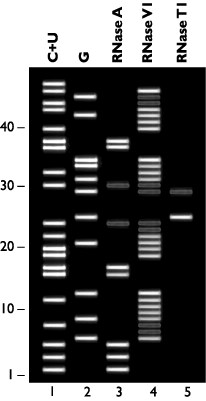
Figure 1. Hypothetical Nuclease Digestion Data from the "Test" RNA Sequence. This "gel" shows the cleavage pattern that would be expected if the "test" RNA were 5' radiolabeled, digested with nucleases in the included Structure Buffer, and then separated by denaturing polyacrylamide gel electrophoresis. Lane 1 (C+U) represents cleavage under denaturing conditions by RNase A at C and U residues. Lane 2 (G) shows cleavage under denaturing conditions by RNase T1 at G residues. Lanes 3-5 represent limited cleavage of the folded "test" RNA by the indicated RNases.
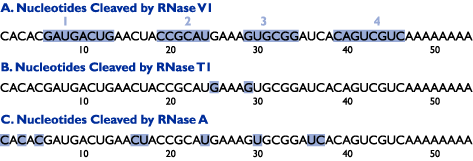
Figure 2. Hypothetical Cleavage Sites of the "Test" RNA. Cleavage sites "generated" by each of the 3 ribonucleases are shaded.
Looking at the hypothetical cleavage pattern "generated" by RNase V1, which preferentially cleaves RNA in double-stranded regions, four regions of RNase V1 cleavage can be identified (Figure 1). Comparing the sequences within these regions (Figure 2, Panel A), two double-stranded regions within the RNA become apparent (Figure 3): regions 1 and 4 form Stem I (8 bp), and regions 2 and 3 form Stem II (6 bp). Less RNase V1 cleavage is seen at residues 7, 8, 23, 24, 29, 30, 44, and 45 (Figure 1), representing the 7:45, 8:44, 23:30, and 24:29 base pairs (Figure 3). This lower reactivity is due to weaker base-pairing of the A:U and G:U base pairs found at these positions.
The RNase T1 cleavage pattern in Figure 1 shows a strong cleavage site at G25. This is the first G in a putative tetraloop structure. A weak cleavage site is seen at G29, indicating that this G has partial single-strand structure. A schematic of the RNase T1 "results" can be seen in Figure 2, Panel B.
RNase A, which cleaves single-stranded RNA at C and U residues, generates eight bands (Figure 1). Two of the bands, representing cleavage at U24 and U30, are relatively weak. Since these two residues are also cleaved by RNase V1, the solution structure is deduced to fluctuate between a tetraloop (25-28) and an 8-base loop (23-30), as the 23:30 and 24:29 base pairs form and melt.
As you can see by this hypothetical example, structure-specific nucleases, when used in conjunction with a structure prediction program like mfold, can provide important structural information about RNA molecules. For more information about analyzing RNA structure, we encourage you to read the references listed below.
Ambion's RNA-Grade RNase A, RNase T1 and RNase V1
Each RNA-Grade ribonuclease is tested for purity to ensure that no contaminating ribonuclease activities are present that could cleave at unanticipated sites. For your convenience, the RNA-Grade RNases are supplied with the following reagents:
- 10X RNA Structure Buffer (1 ml)
- 1X Alkaline Hydrolysis Buffer (1 ml)
- Yeast RNA, 10 mg/ml (100 µl)
- Precipitation/Inactivation Buffer (4.8 ml)
- Gel Loading Buffer II (1.4 ml)
NOTE: RNase T1 is also supplied with 1 ml 1X RNA Sequence Buffer.
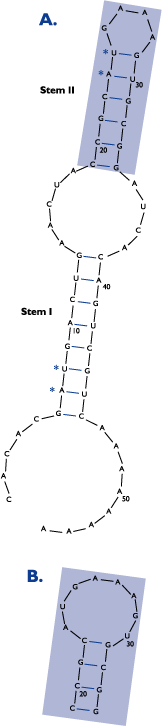
Figure 3. Secondary Structure of the "Test" RNA as Deduced by mfold Analysis. The shaded area represents the loop structures found between nucleotides 23-30. The solution structure in this region is proposed to be a GAAA tetraloop or an 8-base loop (shown in B.). The base pairs marked with asterisks represent regions of weak base pairing within the stem structure.
Selected References on RNA Structure/Function Analysis
1. Moine H, Ehresmann B, Ehresmann C, and Romby P. (1998) Probing RNA structure and function in solution. In: Simons RW and Grunberg-Manago M (editors). RNA Structure and Function, Cold Spring Harbor Laboratory Press. p. 77-115.
A great repository of information on various techniques used for RNA analysis, including enzymatic and chemical interference protocols. Recommended reading for beginners to RNA structure analysis.
2. Knapp G. (1989) Enzymatic approaches to probing of RNA secondary and tertiary structure. Methods Enzymol 180:192-212. Abstract
Detailed protocols for using RNAses in secondary structure analysis. Includes useful information on RNA sequencing and making RNA ladders.
3. Krol A and Carbon P. (1989) A guide for probing native small nuclear RNA and ribonucleoprotein structures. Methods Enzymol 180:212-227. Abstract
Detailed protocols for probing snRNA and snRNP structures. Provides a good description of the mechanisms by which commonly used chemicals (e.g., DMS, ENU, DEPC, and CMCT) modify RNA.
4. Tinoco I Jr, and Bustamante C. (1999) How RNA folds. J Mol Biol 293(2):271-281. Abstract
5. Doudna J A. (2000) Structural genomics of RNA. Nature Struct Biol 7 Suppl:954-956. Abstract
Provides a good perspective on the emerging field of "ribonomics" — structural genomics of RNA.