Search
Unique profiles of miRNA expression
Like mRNAs, miRNA expression profiles appear to vary from tissue to tissue but are similar for identical tissues in different individuals (Lagos-Quintana 2002, Krichevsky 2003, Michael 2003). As would be expected, studies indicate that tissues in developing and mature organisms are characterized by unique profiles of miRNA expression. Comparisons of embryonic stem cells and differentiated cells revealed that miRNA expression profiles change during differentiation (Houbaviy 2003). Notably, at least four miRNAs are expressed at relatively high levels prior to differentiation but are not expressed at all in differentiated cells.
The mirVana™ miRNA Probe Set and mirVana™ miRNA Labeling Kit were used to compare the miRNA expression profiles of 25 human tissues (Figure 4). As expected, miRNA expression profiles vary from tissue to tissue (Lagos-Quintana 2002, Krichevsky 2003, Michael 2003). Interestingly, miRNA profiles are similar between related tissues and distinct between unrelated tissues. For instance, heart and skeletal muscle profiles are very similar, digestive tract tissues cluster, and reproductive organ tissues are similar (Figure 4). The brain miRNA profile, however, is clearly distinct from the other tissues that were analyzed. Follow-up experiments indicated that brain sub-regions exhibited unique miRNA profiles that were clustered according to the relatedness of the sub-regions. The expression data suggest that miRNAs are important factors in differentiating tissues in adult organisms.
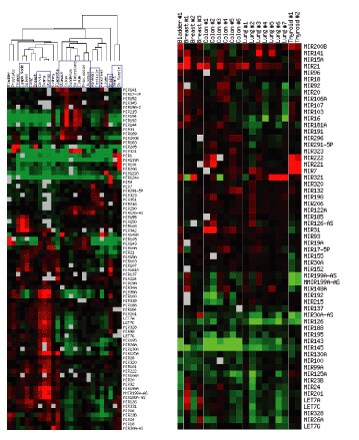
Figure 4 . Data Obtained with the miRNA Expression System
The miRNA expression profiles (y axis) of 20-25 different human normal tissues (x axis) were compared to a pool of all samples in the experiment. Green in the heat map shows miRNAs that are down-regulated in the sample relative to the pool, and red shows miRNAs that are up-regulated in the sample relative to the pool.
The mirVana miRNA Probe Set and mirVana miRNA Labeling Kit were also used to compare the expression profiles of tumor and non-tumor samples from individual cancer patients. Interestingly, a number of miRNAs appear to be routinely under- or over-expressed in tumors. For instance miR-126, miR-143, and miR-145 were expressed at significantly lower levels in more than 80% of the tumor samples compared to their associated normal tissues. miR-21 was found to be over-expressed in 80% of the tumor samples. These miRNAs likely represent biomolecules that directly or indirectly influence oncogenesis. In addition, several miRNAs were found to be differentially expressed in specific types of cancers, suggesting that there are disease-specific miRNAs.
These studies are significant because they show that tissue and cell samples can be defined by their miRNA expression profiles. Categorizing miRNAs based on their co-regulation, or identifying miRNAs that are differentially expressed between different tissue or cell samples will enhance our understanding of miRNA and protein regulation and function. If miRNAs are involved in oncogenesis, inflammatory response, or other disease states, comparative miRNA expression studies might also reveal diagnostic markers or even therapeutic targets. Once the mRNA targets of miRNAs are identified, combining mRNA and miRNA expression profiles will provide a snapshot of genes being regulated at the transcriptional and translational levels and will suggest a more comprehensive list of genes that are important to the biological process being studied.
miRNA Target Sites
The expression of a large number of the predicted 200–300 human miRNA genes (this corresponds to 1% of the protein coding genes) has been confirmed, but the predicted miRNA targets remain to be identified and verified. Several groups have developed algorithms for identification of mRNA sequences that could serve as target sites for known miRNAs. These algorithms take advantage of the observation that each of the known miRNA target sites in animals has perfect or near perfect homology with the first eight bases of the miRNA (Lai 2002). Since each of the known miRNA target sites are in the 3' UTR, algorithms also restrict sequence searches to the 3' UTRs of mRNAs. Another extremely important observation is that homology searches between two or more animal genomes with shared miRNAs confirms that miRNA target sites are conserved. This is a key way to narrow the number of putative target sites. Given that most of the miRNA sequences are conserved between organisms, one would expect that the miRNA target sites would likewise be maintained even though they lie in the typically poorly conserved 3' UTRs of genes. As more is learned about the mRNA targets of the different miRNAs, it will be possible to more accurately assess gene expression for a given sample by combining the profiles of mRNA and miRNA expression.
References
Houbaviy HB, Murray MF Sharp PA (2003) Embryonic stem cell-specific miRNA. Developmental Cell 5: 351-358.
Krichevsky AM, King KS, Donahue CP, Khrapko K, Kosik KS (2003) A miRNA array reveals extensive regulation of miRNAs during brain development. RNA 9: 1274-1281.
Lagos-Quintana M, Rauhut R, Yalcin A, Meyer J, Lendeckel W, Tuschl T (2002) Identification of tissue-specific miRNAs from mouse. Current Biology 12: 735-739.
Michael MZ, O’Connor SM, van Holst Pellekaan NG, Young GP, James RJ (2003) Reduced accumulation of specific microRNAs in colorectal neoplasia. Molecular Cancer Research 1: 882-91.