Search
microRNAs: Definition & Overview
Virtually all of the miRNAs that have been studied in animals reduce steady state protein levels for the targeted gene(s) without impacting the corresponding levels of mRNA (Olsen 1999). The mechanism by which miRNAs reduce protein levels is not fully understood, but one study involving the C. elegans lin-4 miRNA/lin-14 mRNA pair indicates that lin-4 miRNA does not affect the poly(A) tail length, transport to the cytoplasm, nor entry into polysomes of the lin-14 mRNA (Olsen, 1999). If this observation holds true for all animal miRNAs, then downstream steps such as translational elongation, translational termination, or protein stability are likely influenced by miRNAs. Mounting evidence suggests that miRNAs function via a similar enzyme complex as siRNAs. This evidence is summarized below:
- The let-7 miRNA can associate with the RISC in vitro. This indicates that the structure of miRNA does not preclude it from entering the same complex as siRNAs (Hutvagner 2002).
- Endogenous miRNAs can cause degradation of recombinant mRNAs with binding sites that are perfectly complementary to the expressed miRNA (Hutvagner 2002, Zeng 2003). This implies that natural miRNAs associate with RISC in cells. The key feature that distinguishes an miRNA from an siRNA is non-complementarity between the center of the miRNA and the targeted mRNA (Doench 2003, Zeng 2002).
- Immunoprecipitation experiments with antibodies targeting known members of the RISC (eIF2C2, Gemin 3, and Gemin 4) recover endogenous let-7 (Hutvagner 2002), providing further evidence that miRNAs are indeed associated with RISC in cells.
Plant miRNAs differ from animal miRNAs in that many plant miRNAs have perfect homology to their target mRNAs, and they act through the RNAi pathway to cause mRNA degradation (Rhoades 2002). It is likely, however, that some plant miRNAs base-pair imperfectly with their miRNA target sites and act via a pathway similar to animal miRNAs (Figure 3). In plants and yeast there is also evidence that miRNAs are involved in repression of transcription by guiding chromatin methylation.
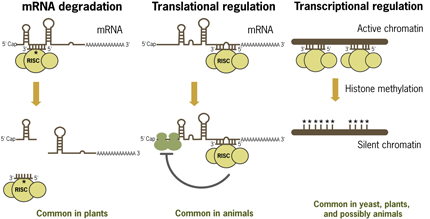
Figure 3 . Mode of Action of miRNAs in Plants and Animals
miRNAs Often Do Not Act Alone
A key observation made by two laboratories is that mRNAs containing multiple, non-overlapping miRNA binding sites are more responsive to miRNA-induced translational repression than those containing a single miRNA binding site (Doench 2003, Zeng 2003). Furthermore, comparisons of repression by miRNAs bound to 2, 4, and 6 binding sites on a reporter construct indicate that translation decreases with each additional site (Zeng 2003). This suggests that the expression of miRNA target genes can be fine-tuned in animals (and potentially plants) by altering the concentrations or identities of miRNAs within cells. This observation coupled with the predictions that many mRNAs have target sites for many different miRNAs suggests that gene expression in various tissues and cells can be greatly influenced by the miRNA populations in those cells. This could also explain why at least some miRNAs have such broad functionality, and conversely why translational control of some genes is so complex. If miRNAs indeed regulate the translation of, but not the stability of target mRNAs, this might at least partially explain why gene expression profiles based on mRNA analysis do not always correlate with protein expression data (Kern 2003).
References
Doench JG, Petersen CP, Sharp PA. (2003) siRNAs can function as miRNAs. Genes Dev 17: 438-42.
Hutvagner G, Zamore PD (2002) A miRNA in a multiple-turnover RNAi enzyme complex. Science 297: 2056-2060.
Kern W, Kohlmann A, Wuchter C, Schnittger S, Schoch C, Mergenthaler S, Ratei R, Ludwig WD, Hiddemann W, Haferlach T. (2003) Correlation of protein expression and gene expression in acute leukemia. Cytometry 55B: 29-36.
Olsen, P. H. and Ambros, V. (1999) The lin-4 regulatory RNA controls developmental timing in C. elegans by blocking LIN-14 protein synthesis after the initiation of translation. Develop Biol 216: 671-680.
Rhoades MW, Reinhart BJ, Lim LP, Burge CB, Bartel B, Bartel DP. (2002) Prediction of plant microRNA targets. Cell 110: 513-20.
Zeng Y, Wagner EJ, Cullen BR (2002) Both Natural and Designed Micro RNAs Can Inhibit the Expression of Cognate mRNAs When Expressed in Human Cells. Molecular Cell 9: 1327-1333.
Zeng Y, Yi R, Cullen BR (2003) miRNAs and small interfering RNAs can inhibit mRNA expression by similar mechanisms. Proc Nat Acad Sci 100: 9779-9784.