Search
Simultaneous Detection of Multiple mRNA Targets with Ribonuclease Protection Assays
The ribonuclease protection assay (RPA) is an extremely sensitive method for detecting and quantitating RNAs (usually mRNAs) in a complex mixture of total cellular RNA (Melton et.al., 1984). The assay is at least 10 times as sensitive as standard Northern blotting, and it enables the investigator to use multiple probes in a single assay. In addition, the RPA is tolerant of partially degraded RNA, allows the use of sample sizes of up to 100 µg in order to further increase sensitivity, and can be used to perform mapping studies such as determining transcript initiation and termination sites and intron-exon boundaries. RPAs can also be used to discriminate between related mRNAs of similar size, which would migrate at similar positions on a Northern blot.
Background
To perform RPAs, an excess of one or more radioactively labeled RNA probes is first hybridized in solution with an RNA sample containing the target RNA. Hybridization is normally an overnight incubation. After the hybridization step, excess unhybridized single-stranded probe molecules are degraded by treatment with RNase. Probe molecules that are hybridized to target molecules, and thus are double-stranded, are protected from the action of the single-strand-specific RNase. In standard procedures, the reaction is then treated with Proteinase K/SDS to inactivate the RNase, followed by phenol/CHCl3 extraction and transfer of the aqueous phase to a clean tube. The protected RNA fragments are then precipitated, resolved on a PAGE gel, and detected by autoradiography with X-ray film. See Figure 1 for a schematic of the RPA procedure.
In 1990, Ambion introduced the RPA™ Kit, the first commercially available ribonuclease protection assay kit. Having all the controls and optimized reagents available in kit form makes the technique more convenient and accessible to a wider range of researchers. Subsequently, Ambion improved this assay by developing a patented reagent, which simultaneously inactivates the RNase and precipitates the protected RNA fragments in a single step. This eliminates the Proteinase K/SDS digestion, phenol:chloroform extraction, and ethanol precipitation steps, and streamlines the procedure to a single tube assay. This technology is incorporated in Ambion's RPA II™ and RPA III™ ribonuclease protection assay Kits, which contain sufficient reagents to perform 100 assays, a positive control probe template and target mRNA, and comprehensive instruction manuals.
In 1990, Ambion introduced the RPA™ Kit, the first commercially available ribonuclease protection assay kit. Having all the controls and optimized reagents available in kit form makes the technique more convenient and accessible to a wider range of researchers. Subsequently, Ambion improved this assay by developing a patented reagent, which simultaneously inactivates the RNase and precipitates the protected RNA fragments in a single step. This eliminates the Proteinase K/SDS digestion, phenol:chloroform extraction, and ethanol precipitation steps, and streamlines the procedure to a single tube assay. This technology is incorporated in Ambion's RPA II™ and RPA III™ ribonuclease protection assay Kits, which contain sufficient reagents to perform 100 assays, a positive control probe template and target mRNA, and comprehensive instruction manuals.
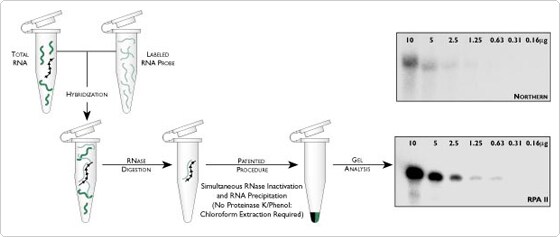
Sensitivity
In Northern blot procedures, some target RNA may remain in the gel or be lost during the transfer steps. More importantly, a fraction of the target RNA transferred is inaccessible to probe because of the manner in which it is bound to the solid support (nylon or nitrocellulose filter). In an RPA, most of the target RNA is available for hybridization to the probe, since it is a true solution hybridization reaction. A second factor contributing to increased sensitivity is that RPAs have lower levels of background signal. With Northerns, signal due to non-specific binding of probe to the filter forms a background against which the target signal must be detected. In the RPA, any non-specific hybrids are digested into small fragments, which are efficiently separated from the protected fragments during electrophoresis. Sensitivity can be further increased by using RNA sample sizes greater than the 10 µg amounts typically used in Northern assays. With RPAs, up to 100 µg of RNA can be used in a single assay. Thus, under these conditions, an RPA can be 100 times more sensitive than a standard Northern (see Figure 1).
Tolerance for Partially Degraded Sample RNA
With Northern blot assays, any degradation of the target mRNA results in loss of signal from the band of interest and the appearance of a smear of smaller molecular weight signal on the autoradiogram. In contrast, in RPAs, the size of the band on the autoradiograph is determined by the size of the probe, which is generally a short (200-500 bases) subregion of the mRNA being detected. Any degradation in the target mRNA which occurs outside the region covered by the probe will not affect the assay. This is especially important with large mRNAs that may be difficult to obtain completely intact. For example, with a 5 kb mRNA analyzed on a Northern, if 20% of the target molecules had a single, random break, there would be a loss of 20% of the signal. With an RPA using a 250 base probe and the same RNA sample, only 1% of the target mRNAs would have a break in the region spanned by the probe.
Simultaneous Detection of Multiple mRNA Targets
In order to measure the level of more than one RNA with standard Northerns, it is generally necessary to go through a repeated process of probing, exposing, stripping and reprobing the blot. In addition to being time consuming, this can be an inconsistent process due to loss of target RNA and incomplete stripping of probe. In contrast, RPAs allow multiple mRNA targets to be assessed simultaneously. This is done by adding multiple probes to the hybridization reaction. These probes must differ in size and each should give a single band when used in an assay by itself. While it is possible to use a dozen probes simultaneously (see below), 5 or 6 is probably a reasonable number if you are designing the probes yourself (Figure 2).
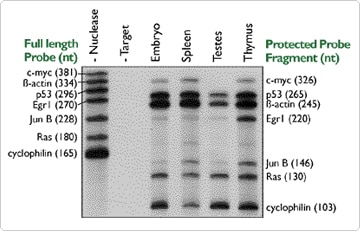
PharMingen has developed sets of ten to twelve probes templates for simultaneous use in RPAs. The probes are co-transcribed from the template set, column purified (no gel purification is used) and co-hybridized to an RNA sample. Figure 3 demonstates the successful use of these template sets in an RPA for simultaneous detection of multiple mRNA targets. In this example, an mApo-3 PharMingen template set was used in conjunction with Ambion's RPA III and MAXIscript™ transcription kits.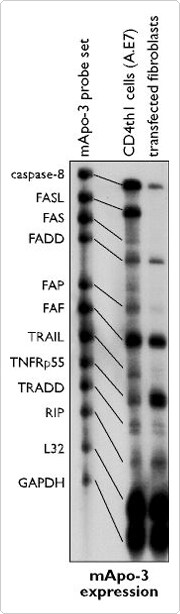
Figure 3. Selective Induction of Apoptosis in Mature Lymphocytes. A.E7 cells (CD4 + T-cell clone) were stimulated with 5 µM of pigeon cytochrome c in the presence of irradiated splenocytes from B10.A mouse strain. After 48 hrs, cells were transferred to fresh medium containing IL-2 and incubated for an additional 48-96 hrs to enerate cycling A.E7 cells that are used in this assay. The fibroblast cells are transfectants expressing I-Ek, ICAM-1 and B7.1 molecules. Total RNA was isolated from the T-cell clone and the transfected fibroblast cells, and 10 µg was used in the RPA III™ assay with 32P-labeled mApo-3 probes (PharMingen) synthesized using Ambion's T7 MAXIscript in Vitro Transcription Kit. 8 x 104 cpm of probe were used per 10 µg of sample RNA.
Lane 1: mApo-3 probe alone; Lane 2: mApo-3 probe + 10 µg CD4+h1 total RNA + RNase; lane 3: mApo-3 probe + 10 µg mouse fibroblast total RNA + RNase.
Lane 1: mApo-3 probe alone; Lane 2: mApo-3 probe + 10 µg CD4+h1 total RNA + RNase; lane 3: mApo-3 probe + 10 µg mouse fibroblast total RNA + RNase.
Using multiple probes clearly increases the quantity of data that can be gathered, but more importantly, increases the quality of the data that is obtained compared to analyzing mRNAs separately. Experimental variability is reduced or eliminated by measuring levels of multiple target mRNAs in the same sample, while one of the probes serves as an internal control by probing an RNA the level of which remains constant (e.g. glyceraldehyde-3-phosphate dehydrogenase (GAPDH), ß-actin, or ribosomal RNA). Collectively, these advantages make RPAs the method of choice for detecting and quantitating multiple mRNAs.
Conclusions
The RPA is a well-proven, extremely sensitive technique for detecting and quantitating even low-abundance RNA species in a complex mixture of total cellular RNA. Multiple probes may be employed in a single hybridization reaction, along with internal and external controls, providing routine detection of as little as 5 femtograms of one or more target mRNA sequences and significant reduction in sample to sample variability. A commercially available kit has made the technique more convenient and accessible to a wider group of researchers.
References
- Melton, D.A., Krieg, P.A., Rebagliati, M.R., Maniatis, T., Zinn, K., and Green, M.R. (1984) Efficient In Vitro Synthesis of Biologically Active RNA and RNA Hybridization Probes From Plasmids Containing a Bacteriophage SP6 Promoter. Nuc. Acids Res. 12: 7035-7056.
- Ngai, J., Dowling, M.M., Buck, L., Axel, R., and Chess, A. (1993) The Family of Genes Encoding Odorant Receptors in the Channel Catfish. Cell 72: 657-666.