Search
The superoxide anion reactive oxygen species (ROS) is generated as a by-product of mitochondrial oxidative phosphorylation. Generation of superoxide, particularly as a result of inhibition of OxPhos Complex I (NADH-quinone oxidoreductase), is a pervasive source of toxicity and mitochondrial dysfunction. The enzyme superoxide dismutase (SOD) performs the key function of counteracting excessive levels of superoxide by converting it to less-reactive hydrogen peroxide. Many of the applications of MitoSOX™ Red indicator for specific detection of mitochondrial superoxide in live cells (Figure 1) and tissues discussed in this article revolve around SOD and Complex I, disorders in their expression, structure, and function, and their ultimate consequences in pathologies including amyotrophic lateral sclerosis (ALS), Parkinson’s disease, and Alzheimer’s disease [1,2]. Other applications described here include cardiovascular biology and oxidative damage studies, as well as examples of the probe’s utility in both microscopy and flow cytometry.
 | Figure 1. Detecting superoxide in live cells using MitoSOX™ Red superoxide indicator. Live 3T3 fibroblasts were treated with iron porphyrin FeTCPP, a superoxide dismutase (SOD) mimetic (Top), or left untreated (Bottom). Cells were then labeled with MitoSOX™ Red indicator in combination with blue-fluorescent Hoechst 33342 nuclear stain. Knockdown of the mitochondrial fluorescence signal by SOD mimetics such as FeTCPP and the manganese porphyrin MnTBAP provides valuable negative controls in MitoSOX™ Red superoxide detection assays. Useful positive controls (not shown) include treatment with antimycin A, doxorubicin, or high glucose. |
Discriminate Superoxide From Other ROS
MitoSOX™ Red indicator is a derivative of dihydroethidium (also known as hydroethidine, HE; Figure 2). The cationic triphenylphosphonium substituent is responsible for the electrophoretically driven uptake of the probe in actively respiring mitochondria. Oxidation of MitoSOX™ Red indicator (or HE) by superoxide results in hydroxylation at the 2-position (Figure 2). 2-hydroxyethidium (and the corresponding derivative of MitoSOX™ Red indicator) exhibits a fluorescence excitation peak at ~400 nm [3] that is absent in the excitation spectrum of the ethidium oxidation product generated by ROS other than superoxide (Figure 3). Thus, fluorescence excitation at 400 nm with emission detection at ~590 nm provides optimum discrimination of superoxide from other ROS [3–5].
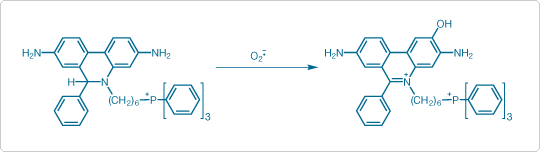
Figure 2. Oxidation of MitoSOX™ Red indicator to 2-hydroxy-5-(triphenylphosphonium)hexylethidium by superoxide (O2 –.)

Figure 3. Fluorescence excitation and emission spectra of 2-hydroxyethidium bound to DNA. The excitation peak at ~400 nm that distinguishes 2-hydroxyethidium from ethidium is marked with an arrow. Click image to enlarge
Neuroscience Applications
MitoSOX™ Red indicator has proven useful for applications in both neurons and glial cells. In many of these investigations, MitoSOX™ Red indicator is just one of several probes used to provide a correlative assessment of multiple physiological parameters. For example, in mouse cortical neurons expressing caspase-cleaved tau microtubule associated protein, mitochondrial superoxide generation (measured using MitoSOX™ Red indicator) has been correlated with cytosolic and mitochondrial calcium (measured using fluo-3, AM and rhod-2, AM, respectively) and mitochondrial membrane potential (measured using TMRM) [6].
The relationship of mitochondrial superoxide generation to dopamine transporter activity (measured using the aminostyryl dye substrate 4-Di-1-ASP) has been investigated in mouse brain astrocytes [7]. In PTEN-induced kinase 1 (PINK1) knockout mouse neurons, elevations in both mitochondrial and cytosolic superoxide (measured using MitoSOX™ Red indicator and HE, respectively) are associated with impaired glucose uptake (measured using 2-NBDG) and mitochondrial calcium overload (monitored using rhod-FF, AM) [8].
Cardiovascular Biology Applications
The cardiovascular system, like the nervous system, is a major bioenergetic consumer and is therefore another arena in which mitochondrial dysfunction assumes critical importance. MitoSOX™ Red indicator has been used for confocal microscopy analysis of ROS production by mitochondrial NO synthase (mtNOS) in permeabilized feline ventricular myocytes [9] and, in combination with Amplex® Red Reagent, for measurement of mitochondrial superoxide and hydrogen peroxide production in rat vascular endothelial cells [10]. Mitochondrial dysfunction plays a central role in the development of diabetes complications [11], leading to interest in characterizing mitochondrial membrane potential and superoxide generation responses to elevated saturated fatty acid levels in cardiomyocytes from normal and obese insulin-resistant mice using TMRE and MitoSOX™ Red indicator [12].
Beyond the Microscope: Applications In Flow Cytometry
In addition to imaging and microscope photometry measurements, several flow cytometry applications of MitoSOX™ Red indicator have also been reported. These include flow cytometric analyses in intact mouse beta cells and isolated mitochondria [13], human colon, pancreatic, breast, liver, and lung carcinoma cells [14], myelogenous leukemia cell lines HL-60 and KG1a [15], and rat primary astrocytes [16]. Detailed protocols for simultaneous measurements of mitochondrial superoxide generation and apoptotic markers (using allophycocyanin–annexin V and SYTOX® Green Stain) in human coronary artery endothelial cells by flow cytometry have been published by Mukhopadhyay and coworkers [17]. Detection of MitoSOX™ Red indicator by flow cytometry typically uses the FL2 emission channel (585/42 nm).
Oxidative Damage Applications
A common application of MitoSOX™ Red indicator is the correlation of superoxide generation with the accumulation of oxidative damage products such as lipid peroxides and modified DNA bases such as 8-oxodG. Mitochondrial DNA (mtDNA) is located in close proximity to the electron transport chain complexes and is consequently more susceptible to oxidative damage than nuclear DNA. However, superoxide exposure (induced by rotenone inhibition of Complex I and detected by MitoSOX™ Red indicator) does not result in significant mtDNA mutagenesis (assessed by Southern blotting and DNA sequencing), due to enhanced mtDNA degradation that prevents the accumulation of mutagenic lesions [18].
Analysis of mitochondrial superoxide using MitoSOX™ Red indicator and lipid peroxidation using BODIPY® 581/591 C11 has facilitated investigation of oxidative stress generation mechanisms in human spermatozoa [19]. Oxidative damage products may have compounding effects by instigating further ROS generation. Using quantitative imaging of MitoSOX™ Red indicator, researchers from the University of Manitoba found that treatment of vascular endothelial cells with extensively oxidized low-density lipoprotein (LDL) resulted in near doubling of mitochondrial superoxide generation compared to treatment with normal LDL [20].
A Versatile Probe For ROS Studies
Using MitoSOX™ Red indicator for specific detection of superoxide in live cells and tissues has proven to be valuable in a variety of applications and research areas, from the pathogenesis of neurodegenerative disease to the development of cardiovascular problems.
- Zimmerman MC, Oberley LW, Flanagan SW (2007) J Neurochem 102:609−618.
- Marella M, Seo BB, Matsuno-Yagi A et al. (2007) J Biol Chem 282:24146–24156.
- Zielonka J, Vasquez-Vivar J, Kalyanaraman B (2008) Nat Protoc 3:8–21.
- Robinson KM, Janes MS, Pehar M et al. (2006) Proc Natl Acad Sci U S A 103:15038–15043.
- Robinson KM, Janes MS, Beckman JS (2008) Nat Protoc 3:941–947.
- Quintanilla RA, Matthews-Roberson TA, Dolan PJ et al. (2009) J Biol Chem 284:18754–18766.
- Mallajosyula JK, Kaur D, Chinta SJ et al. (2008) PLoS ONE 3:e1616.
- Gandhi S, Wood-Kaczmar A, Yao Z et al. (2009) Mol Cell 33:627–638.
- Dedkova EN, Blatter LA (2009) J Physiol 587:851–872.
- Ungvari Z, Labinskyy N, Gupte S et al. (2008) Am J Physiol Heart Circ Physiol 294:H2121–H2128.
- Quijano C, Castro L, Peluffo G et al. (2007) Am J Physiol Heart Circ Physiol 293:H3404−H3414.
- Fauconnier J, Andersson DC, Zhang SJ et al. (2007) Diabetes 56:1136–1142.
- Gusdon AM, Chen J, Votyakova TV et al. (2009) Methods Enzymol 456:439–457.
- Zhao W, Mackenzie GG, Murray OT et al. (2009) Carcinogenesis 30:512−519.
- Davies NJ, Hayden RE, Simpson PJ et al. (2009) Cancer Res 69:4769–4775.
- Cassina P, Cassina A, Pehar M et al. (2008) J Neurosci 28:4115–4122.
- Mukhopadhyay P, Rajesh M, Haskó G et al. (2007) Nat Protoc 2:2295−2301.
- Shokolenko I, Venediktova N, Bochkareva A et al. (2009) Nucleic Acids Res 37:2539–2548.
- Koppers AJ, De Iuliis GN, Finnie JM et al. (2008) J Clin Endocrinol Metab 93:3199–3207.
- Roy Chowdhury SK, Sangle GV, Xie X et al. (2010) Am J Physiol Endocrinol Metab 298:E89–E98.
For Research Use Only. Not intended for any animal or human therapeutic or diagnostic use.
Mitochondrial Biology Poster
 | Probes for Mitochondrial Morphology and Function |
Learn More
Find more information on autophagy-related reagents.
Download This Article
Quick Product View
See a complete listing of the products discussed in this article.
View products
