Search

Related Product Information
Introduction
The GeneSwitch™ System Overview
The GeneSwitch™ System is a mifepristone-inducible mammalian expression system originally developed by Wang et al. (1994) that uses a combination of regulatory components derived from various human and yeast transcription factors. Mifepristone regulation in the GeneSwitch™ System is based on an autoregulatory feedback loop that involves the binding of a GAL4 regulatory fusion protein (see below) to GAL4 upstream activating sequences (UAS) in both the promoter controlling expression of the GAL4 regulatory fusion protein and the promoter controlling expression of the gene of interest. The major components of the GeneSwitch™ System include:
- An inducible expression plasmid, pGene/V5-His, for expression of your gene of interest under the control of a hybrid promoter containing GAL4 upstream activating sequences (UAS) and the adenovirus E1b TATA box
- A regulatory plasmid, pSwitch, which encodes a fusion protein consisting of the yeast GAL4 DNA binding domain (DBD), a truncated human progesterone receptor ligand binding domain (hPR-LBD), and the human p65 activation domain (AD) from NF-kB
- A control expression plasmid containing the lacZ gene, pGene/V5-His/lacZ, which when cotransfected with pSwitch, expresses beta-galactosidase upon induction with mifepristone
- Mifepristone for inducing expression
Description of the GeneSwitch™ System
The GeneSwitch™ System exploits the fact that transcription factors are composed of functional domains (e.g. DNA binding domain (DBD) or activation domain (AD)). In the GeneSwitch™ System, a hybrid regulatory protein containing a DBD from the yeast GAL4 protein, a truncated ligand binding domain (LBD) from the human progesterone receptor, and an AD from the human NF-kB protein is expressed. This hybrid regulatory protein binds to the synthetic steroid, mifepristone, and functions as a ligand-dependent transcription factor to induce expression of the gene of interest as well as its own expression.
The first major component of the GeneSwitch™ System is the pGene/V5-His inducible expression plasmid. Expression of your gene of interest from the inducible expression plasmid is controlled by a hybrid promoter consisting of Saccharomyces cerevisiae GAL4 upstream activating sequences (UAS) (Giniger et al., 1985; Wang et al., 1994) linked to the TATA box sequence from the adenovirus major late E1b gene (Lillie and Green, 1989). Contained within the GAL4 UAS are 6 copies of the 17 nucleotide sequence, 5' -(T/C)GGAGTACTGTCCTCCG-3' that constitute the binding site for the yeast GAL4 transcription factor. Each 17 nucleotide sequence serves as the binding site for two molecules of the GAL4 DBD (Marmorstein et al., 1992).
The second major component of the GeneSwitch™ System is the pSwitch regulatory vector which expresses the GAL4-DBD/hPR-LBD/p65-AD regulatory fusion protein under the control of a hybrid promoter consisting of a GAL4 UAS (containing 4 copies of the GAL4 binding site) linked to a minimal promoter from the Herpes Simplex Virus thymidine kinase (TK) gene.
Mechanism of Autoregulation and Induction
The pSwitch regulatory vector and your pGene/V5-His construct will be introduced into the mammalian cell line of choice by cotransfection. In the absence of mifepristone, low basal transcription of the GAL4-DBD/hPR-LBD/p65-AD fusion gene from pSwitch occurs from the minimal thymidine kinase (TK) promoter. Once translated into protein, the GAL4-DBD/hPR-LBD/p65-AD regulatory protein (GeneSwitch™ protein) is predominantly localized in the nucleus in an inactive form. Upon addition, mifepristone binds with high affinity to the truncated hPR-LBD in the GeneSwitch™ protein (Kd ~ 3 x 10-9 M) (Vegeto et al., 1992) and causes a conformational change in the hPR-LBD, resulting in dimerization of the GeneSwitch™ protein and conversion to an active form. The ligand-bound GeneSwitch™ homodimer then interacts with the GAL4 binding sites in the GAL4 UAS of both pGene/V5-His and pSwitch and activates transcription of both the gene of interest from the E1b TATA box and the regulatory fusion gene from the minimal TK promoter. Expression of the gene of interest is further amplified through initiation of an autoregulatory feedback loop whereby the ligand-bound GeneSwitch™ protein upregulates expression of its own gene. Newly synthesized GeneSwitch™ protein binds to mifepristone, undergoes a conformational change to an active state, and induces transcription of the gene of interest as well as its own gene. . For more information about the progesterone receptor, its mechanism of action, and the ligand-receptor interaction, refer to published reviews and articles (DeFranco, 1998; Gasc et al., 1989; Guiochon-Mantel et al., 1989; Simons, 1998; Ylikomi et al., 1992).
Website
The GeneSwitch™ System is a modified version of the gene switch regulatory system originally described by Wang, et al. (1994). For additional information and publications relating to the gene switch technology, refer to the website administered by Valentis, Inc.
For specific information pertaining to the GeneSwitch™ System, download the user manual or call Technical Service.
The GAL4-DBD/hPR-LBD/p65-AD Regulatory Protein
The GAL4-DBD/hPR-LBD/p650-AD regulatory fusion protein (GeneSwitch™ protein) expressed from the pSwitch plasmid is a 73 kDa hybrid protein composed of the specified regulatory domains from the three transcription factors listed below.
Transcription Factor | Regulatory Domain | Benefit of Regulatory Domain | References |
Saccharomyces cerevisiae GAL4
|
DNA binding domain (DBD)
Amino acids 1-93 in native protein
|
Allows GeneSwitch
™ protein to bind to DNA via GAL4 binding sites in the GAL4 UAS
|
(Giniger
et al., 1985; Laughon and Gesteland, 1984; Marmorstein
et al., 1992)
|
Human progesterone receptor (hPR)
|
Truncated ligand binding domain (LBD)
Amino acids 640-914 in native protein
|
Allows mifepristone-specific binding and activation of the GeneSwitch
™ protein
|
(Kastner
et al., 1990; Misrahi
et al., 1987; Vegeto
et al., 1992; Wang
et al., 1997)
|
Human p65 subunit of NF-kB
|
Activation domain (AD)
Amino acids 283-551 in native protein
|
Permits activation of gene transcription
|
(Burcin
et al., 1999; Deloukas and Loon, 1993; Ruben
et al., 1991; Schmitz and Baeuerle, 1991)
|
- In the GeneSwitch™ System, the GeneSwitch™ protein functions as a ligand-dependent transcription factor to activate expression of both the gene of interest and its own gene via binding to GAL4 sites within the GAL4 UAS. The combination of functional domains from the transcription factors described above allows the GeneSwitch™ protein to exhibit the following characteristics. Because the GAL4 DBD is derived from a yeast protein, the GeneSwitch™ protein has no effect on endogenous genes and can only activate transcription of genes whose expression is controlled by a GAL4 UAS (i.e. the gene of interest and the regulatory fusion gene).
- The GAL4 DBD binds to an individual 17 nucleotide GAL4 binding site as a homodimer (Carey et al., 1989; Marmorstein et al., 1992). The pGene/V5-His and pSwitch plasmids contain 6 and 4 copies of the GAL4 binding site, respectively, although it is not known if all of the GAL4 binding sites are occupied at any given time. The truncated hPR-LBD contains a 19 amino acid deletion from its C-terminal end that abolishes its ability to bind to progesterone, other endogenous steroid hormones, or other progesterone agonists, but still enables it to bind with high affinity to mifepristone (Vegeto et al., 1992; Wang et al., 1994; Wang et al., 1997).
- The p65 AD is a strong transcriptional activator but is derived from a human protein to minimize possible toxic or pleiotrophic effects associated with viral transactivation domains (Abruzzese et al., 1999; Burcin et al., 1999)
Hallmarks of Mifepristone Induction
The synthetic steroid, mifepristone, is used as an inducing agent in the GeneSwitch™ System. The hallmarks of mifepristone induction in this system are listed below:
- Induction of gene expression requires extremely low concentrations of mifepristone (i.e. 1 x 10-8 M mifepristone)
- The concentration of mifepristone required for half-maximal induction in the GeneSwitch™ System is approximately 10-10 M
- The concentrations of mifepristone used for gene induction exert no known toxic or pleiotrophic effects on mammalian cells that lack endogenous progesterone and glucocorticoid receptors (Vegeto et al., 1992; Wang et al., 1994)
Experimental Outline
To express your gene of interest using the GeneSwitch™ System, you will perform the following steps: (see below for a diagram):
- Ligate your gene of interest into the multiple cloning site of the pGene/V5-His inducible expression vector.
- Cotransfect your pGene/V5-His construct and the pSwitch regulatory plasmid into the mammalian cell line of choice.
- Add mifepristone to induce expression of the gene of interest via an autoregulatory feedback loop involving the GAL4-DBD/hPR-LBD/p65-AD regulatory fusion protein.
- Assay for expression of your recombinant protein of interest.
Note: The positive control vector containing the lacZ gene (pGene/V5-His/lacZ) can be transiently cotransfected into mammalian cells with pSwitch to demonstrate that the system is working properly in your cell line. Stable cell lines expressing the GAL4 DBD/hPR-LBD/p65 AD regulatory fusion protein from pSwitch can be established to serve as hosts for inducible expression vector-based constructs.
The GeneSwitch™ System manual is supplied with the kits listed below. The Core System includes vectors, primers for sequencing, and the inducing agent, mifepristone. The Complete System includes the Core System plus selection agents. See below for a detailed description of the contents of each GeneSwitch™ System.
Kit Contents
Both the GeneSwitch™ Complete and the GeneSwitch™ Core Systems include the following regulatory vector, sequencing primers, and inducing agent. Store all reagents at -20°C.
Reagent | Amount Supplied | Comments |
Zeocin
™ |
1 g
|
Selection agent for the pGene/V5-His plasmid
|
Hygromycin B
|
1 g
|
Selection agent for the pSwitch regulatory plasmid
|
Expression Vectors
Each GeneSwitch™ Complete and Core System also includes an inducible expression vector for cloning your gene of interest and a corresponding positive control vector containing the lacZ gene as described below. The inducible expression vector is supplied in three reading frames to facilitate cloning the gene of interest in frame with a C-terminal peptide containing the V5 epitope and a polyhistidine tag.
Vector Amount
pGene/V5-His A, B, C 20 µg each, lyophilized in TE, pH 8.0
pGene/V5-His/lacZ 20 µg, lyophilized in TE, pH 8.0
Selection Agents
In addition to the vectors and primers provided in the GeneSwitch™ Core System, the GeneSwitch™ Complete System also includes the selection agents listed below. Zeocin™ is supplied in 8 x 1.25 ml aliquots at a concentration of 100 mg/ml.
Reagent | Amount Supplied | Comments |
Zeocin
™ |
1 g
|
Selection agent for the pGene/V5-His plasmid
|
Hygromycin B
|
1 g
|
Selection agent for the pSwitch regulatory plasmid
|
Introduction
This section contains guidelines to propagate the GeneSwitch™ vectors.
E. coli Strain
Many E. coli strains are suitable for the propagation of the GeneSwitch™ vectors including TOP10 (Catalog no. C610-00) and DH5a. We recommend propagating the GeneSwitch™ vectors in recombination deficient (recA) and endonuclease A deficient (endA) E. coli strains. For your convenience, we offer TOP10 and DH5a E. coli as chemically competent or electrocompetent cells.
| Item | Quantity | Catalog no. |
One Shot
® TOP10 Chemically Competent
E. coli |
20 x 50 µl
|
C4040-03
|
One Shot
® TOP10 Electrocomp
™E. coli |
20 x 50 µl
|
C4040-52
|
One Shot
® MAX Efficiency
® DH5™-T1
R Chemically Competent
E. coli |
20 x 50 µl
|
12297-016
|
Transformation Method
You may use any method of choice for transformation. Chemical transformation is the most convenient, but electroporation is the most efficient and the method of choice for large plasmids.
Maintenance of Plasmids
The GeneSwitch™ vectors contain the ampicillin resistance gene to allow selection of the plasmid using ampicillin. The pGene/V5-His A, B, C and pGene/V5-His/lacZ vectors also contain the Zeocin™ resistance gene for selection of the plasmids using Zeocin™. To propagate and maintain the GeneSwitch™ plasmids, use the following procedure:
1. Resuspend each vector in 20 µl sterile water to prepare a 1 µg/µl stock solution. Store the stock solution at -20°C.
2. Use the stock solution to transform a recA, endA E. coli strain like TOP10, DH5a, or equivalent.
3. Select transformants on the appropriate plates as follows:
- For the pGene/V5-His A, B, C and pGene/V5-His/lacZ plasmids, select on LB plates containing 50 to 100 µg/ml ampicillin or 25 to 50 µg/ml Zeocin™ in Low Salt LB
- For the pSwitch plasmid, select on an LB plate containing 50 to 100 µg/ml ampicillin
Note: For fast and easy microwaveable preparation of Low Salt LB agar containing ampicillin or Zeocin™, imMedia™ Amp Agar (Catalog no. Q601-20) or imMedia™ Zeo Agar (Catalog no. Q621-20) are available. For more information, see our website or call Technical Service.
4. Prepare a glycerol stock from each transformant containing plasmid for long-term storage.
Introduction
You will express your gene of interest from the pGene/V5-His inducible expression vector. The pGene/V5-His vector is supplied with the multiple cloning site in three reading frames (A, B, and C) to facilitate cloning your gene of interest in frame with the C-terminal peptide containing the V5 epitope and a polyhistidine (6xHis) tag. Use the diagrams provided to help you design a strategy to clone your gene of interest in frame with the C-terminal peptide. General considerations for cloning and transformation are discussed in this section.
General Molecular Biology Techniques
For assistance with E. coli transformations, restriction enzyme analysis, DNA biochemistry, and plasmid preparation, refer to Molecular Cloning: A Laboratory Manual (Sambrook et al., 1989) or Current Protocols in Molecular Biology (Ausubel et al., 1994).
Cloning Considerations
Your insert should contain a Kozak translation initiation sequence with an ATG start codon for proper initiation of translation (Kozak, 1987; Kozak, 1991; Kozak, 1990). An example of a Kozak consensus sequence is provided below. Please note that other sequences are possible, but the G or A at position –3 and the G at position +4 are the most critical for function (shown in bold). The ATG initiation codon is shown underlined.
(G/A)NNATGG
To express your gene as a recombinant fusion protein, you must clone your gene in frame with the C-terminal peptide. The vector is supplied in three reading frames to facilitate cloning. If you wish to express your protein WITHOUT the C-terminal peptide, be sure to include a stop codon.
Multiple Cloning Site of Version A
Below is the multiple cloning site for pGene/V5-His A. Restriction sites are labeled to indicate the cleavage site. The boxed nucleotides indicate the variable region. Note that there is a stop codon between the Spe I and BstX I sites in the polylinker. Also, note that the sequences of the GAL4 binding sites are not identical. The multiple cloning site has been confirmed by sequencing and functional testing. The complete sequence of pGene/V5-His A is available for download or from Technical Service. 
*Note that there are two BstX I sites in the polylinker.
Multiple Cloning Site of Version B
Below is the multiple cloning site for pGene/V5-His B. Restriction sites are labeled to indicate the cleavage site. The boxed nucleotides indicate the variable region. Note that the sequences of the GAL4 binding sites are not identical. The multiple cloning site has been confirmed by sequencing and functional testing. The complete sequence of pGene/V5-His B is available for download or from Technical Service.
*Note that there are two BstX I sites in the polylinker
Multiple Cloning Site of Version C
Below is the multiple cloning site for pGene/V5-His C. Restriction sites are labeled to indicate the cleavage site. The boxed nucleotides indicate the variable region. Note that the sequences of the GAL4 binding sites are not identical. The multiple cloning site has been confirmed by sequencing and functional testing. The complete sequence of pGene/V5-His C is available for download or from Technical Service.
*Note that there are two BstX I sites in the polylinker.
E. coli Transformation
Transform your ligation mixtures into a competent recA, endA E. coli strain (e.g. TOP10, DH5a) and select on LB agar plates containing 50 to 100 mg/ml ampicillin or 25 to 50 mg/ml Zeocin™ in Low Salt LB. Select 10-20 clones and analyze for the presence and orientation of your insert.
Any E. coli strain that contains the complete Tn5 transposable element (i.e. DH5aF' IQ, SURE, SURE2) encodes the ble (bleomycin) resistance gene. These strains will confer resistance to Zeocin™. For the most efficient selection with Zeocin™, we recommend that you choose an E. coli strain that does not contain the Tn5 gene (i.e. TOP10, TOP10F' ).
Zeocin™ Selection in E. coli
To facilitate selection of Zeocin™-resistant E. coli, the salt concentration of the medium must remain low (< 90 mM) and the pH must be 7.5. Prepare Low Salt LB broth and plates.
Failure to lower the salt content of your LB medium will result in non-selection due to inactivation of the Zeocin™.
We recommend that you sequence your construct with the pGene Forward and BGH Reverse primers provided with the kit to confirm that your gene contains an ATG start codon and is cloned in frame with the C-terminal peptide (if desired). Refer to the diagrams for the sequences and location of the priming sites.
Preparing the Glycerol Stock
Once you have identified the correct clone, be sure to purify the colony and make a glycerol stock for long-term storage. It is also a good idea to keep a DNA stock of your plasmid at -20°C.
- Streak the original colony out on an LB plate containing 50 mg/ml ampicillin or 25-50 mg/ml Zeocin™ in Low Salt LB. Incubate the plate at 37°C overnight.
- Isolate a single colony and inoculate into 1-2 ml of LB containing 50 mg/ml ampicillin or 25-50 mg/ml Zeocin™ in Low Salt LB.
- Grow the culture to mid-log phase (OD600 = 0.5-0.7).
- Mix 0.85 ml of culture with 0.15 ml of sterile glycerol and transfer to a cryovial.
- Store at -80°C.
Introduction
Once you have cloned your gene of interest into pGene/V5-His and have prepared clean plasmid preparations of your pGene/V5-His construct and pSwitch, you are ready to cotransfect the plasmids into the mammalian cell line of choice. We recommend that you include the positive control vector (see below) and a mock transfection to evaluate your results. General guidelines are provided on the next page for cotransfection and induction.
Plasmid Preparation
Plasmid DNA for transfection into eukaryotic cells must be very clean and free from phenol and sodium chloride. Contaminants will kill the cells, and salt will interfere with lipids, decreasing transfection efficiency. We recommend isolating plasmid DNA using the S.N.A.P.™ MiniPrep Kit (10-15 mg DNA, Catalog no. K1900-01), the S.N.A.P.™ MidiPrep Kit (10-200 mg DNA, Catalog no. K1910-01), or CsCl gradient centrifugation.
Methods of Transfection
For established cell lines (e.g. HeLa, COS-1), consult original references or the supplier of your cell line for the optimal method of transfection. We recommend that you follow exactly the protocol for your cell line. Pay particular attention to medium requirements, when to pass the cells, and at what dilution to split the cells. Further information is provided in Current Protocols in Molecular Biology (Ausubel et al., 1994).
Methods for transfection include calcium phosphate (Chen and Okayama, 1987; Wigler et al., 1977), lipid-mediated (Felgner et al., 1989; Felgner and Ringold, 1989) and electroporation (Chu et al., 1987; Shigekawa and Dower, 1988). We offer the Calcium Phosphate Transfection Kit and cationic lipid-based reagents including Lipofectamine™ 2000 for transfection. For more information about the transfection reagents available, visit our website or call Technical Service.
Positive Control
pGene/V5-His/lacZ is provided as a positive control vector for mammalian cell transfection and expression and may be used to optimize induction conditions for your cell line. Cotransfection of the positive control vector and pSwitch results in the induction of beta-galactosidase expression upon addition of mifepristone. A successful cotransfection will result in beta-galactosidase expression that can be easily detected.(see below).
Assay for β-galactosidase Activity
You may assay for beta-galactosidase expression by activity assay using cell-free lysates (Miller, 1972) or by staining the cells for activity. We offer the β-Gal Assay Kit (Catalog no. K1455-01) and the β-Gal Staining Kit (Catalog no. K1465-01) for fast and easy detection of β-galactosidase expression.
Mifepristone
In the GeneSwitch™ System, the synthetic steroid, mifepristone, is used as an inducing agent to activate transcription of the gene of interest as well as transcription of the GeneSwitch™ regulatory protein. Although mifepristone generally behaves as a progesterone antagonist (Philibert et al., 1985), the compound actually functions as an agonist in the GeneSwitch™ System through binding to the truncated hPR-LBD in the GeneSwitch™ protein (Vegeto et al., 1992; Wang et al., 1994; Wang et al., 1997). Once bound to mifepristone, the hPR-LBD undergoes a conformational change that converts the GeneSwitch™ protein from an inactive to an active form. The active regulatory protein then acts as a ligand-dependent transcription factor to activate expression of the gene of interest and its own gene. Extremely low doses of mifepristone (nanomolar range) are required to activate gene transcription.
Note: Mifepristone can bind to progesterone and glucocorticoid receptors in cell lines that contain the native receptors. In these cell lines (e.g. some Chinese Hamster Ovary (CHO) and HeLa cell lines), mifepristone may exert antagonistic effects against the native receptors. However, the concentrations of mifepristone used to induce gene expression in the GeneSwitch™ System exert no known toxic or pleiotrophic effects on mammalian cells that lack endogenous progesterone and glucocorticoid receptors.
Mifepristone is toxic. Do not ingest or inhale the powder or solutions containing the drug. Use caution when handling large quantities of mifepristone. At high doses (> 100 mg), mifepristone may impair fertility and may cause harm to the unborn child. Always wear gloves, a laboratory coat, and safety glasses or goggles when handling mifepristone and mifepristone-containing solutions.
We have found that by varying the ratio of pSwitch and pGene/V5-His plasmids that we transiently cotransfect into mammalian cells, we can minimize basal transcription and optimize mifepristone regulation of the gene of interest from the pGene/V5-His expression plasmid.
In general, we recommend that you cotransfect your mammalian host cell line with a ratio of at least 1:4 (w/w) pSwitch:pGene/V5-His plasmid DNA. Note that results may vary depending on the cell line and the gene of interest; therefore, you may want to empirically determine an optimal ratio of pSwitch and pGene/V5-His to use for cotransfection of your particular cell line.
Although the GeneSwitch™ System is designed for use in a wide range of mammalian cell lines, there may be some host cell lines that are not suitable due to detectable basal transcription or low inducibility of the gene of interest. We have observed detectable basal expression as well as low inducibility of the lacZ gene from pGene/V5-His/lacZ (after cotransfection with pSwitch) in Chinese Hamster Ovary (CHO) cells; therefore, we do not recommend using CHO cells to perform transient transfection experiments with the GeneSwitch™ System.
Cotransfection and Induction with Mifepristone
General guidelines are provided below to cotransfect your pGene/V5-His construct (or the control plasmid) and pSwitch into the mammalian cell line of your choice and to induce expression of your protein of interest with mifepristone. Since every cell line is different and may require a different method of transfection, some empirical experimentation may be needed to determine the optimal conditions for inducible expression. In general:
- Use cells that are approximately 60% confluent for transfection.
- Cotransfect the pSwitch plasmid and your pGene/V5-His construct at a ratio of at least 1:4 (w:w) or at your optimally-determined ratio into the cell line of choice using your preferred method. Absolute amounts of plasmid will vary depending on the method of transfection and the cell line used.
- After transfection, add fresh medium and allow the cells to recover for 24 hours before induction.
- Remove medium and add fresh medium containing the appropriate concentration of mifepristone to the cells. In general, we recommend that you add mifepristone to a final concentration of 1 x 10-8 M (10 ml of a 10 mM stock per 10 ml of medium) to the cells and incubate the cells for 24 hours at 37°C.
Detection of Recombinant Fusion Proteins
You may use our Anti-V5 antibodies or Anti-His (C-term) antibodies to detect expression of your recombinant fusion protein from pGene/V5-His if you have cloned your gene in frame with the C-terminal peptide (see page vii for ordering information). In addition, the Positope™ Control Protein (Catalog no. R900-50) is available for use as a positive control for detection of fusion proteins containing a V5 epitope and a polyhistidine (6xHis) tag. For more information, refer to our website or call Technical Service. To detect your fusion protein by western blot, you will need to prepare a cell lysate from transfected cells. We recommend that you perform a time course to optimize expression of the fusion protein (e.g. 12, 24, 36, 48 hours etc. after mifepristone induction). To assay for beta-galactosidase expression from the pGene/V5-His/lacZ control plasmid, we generally harvest cells after 24 hours of induction with mifepristone. Use the protocol below to lyse cells. Other protocols are suitable.
- Wash cell monolayers (~5 x 105 to 1 x 106 cells) once with phosphate-buffered saline (PBS, Catalog no. 10010-023).
- Scrape cells into 1 ml PBS and pellet the cells at 1500 x g for 5 minutes.
- Resuspend in 50 ml Cell Lysis Buffer. Other cell lysis buffers are suitable. Vortex.
- Incubate cell suspension at 37°C for 10 minutes to lyse the cells. Note: You may prefer to lyse the cells at room temperature or on ice if degradation of your protein is a potential problem.
- Centrifuge the cell lysate at 10,000 x g for 10 minutes at +4°C to pellet nuclei and transfer the supernatant to a fresh tube. Assay the lysate for protein concentration.
- Add SDS-PAGE sample buffer to a final concentration of 1X and boil the sample for 5 minutes.
- Load 20 mg of lysate onto an SDS-PAGE gel and electrophorese. Use the appropriate percentage of acrylamide to resolve your fusion protein.
Note:
The C-terminal peptide containing the V5 epitope and the polyhistidine (6xHis) tag will add approximately 5 kDa to the size of your protein.
Purifying Your Recombinant Protein
You will need approximately 5 x 106 to 1 x 107 transfected cells to purify your protein on a 2 ml ProBond™ or Ni-NTA column. Refer to the procedure on page 21 to prepare cells for lysis.
Optimizing Expression
You may vary the concentration of mifepristone (1 x 10-7 M to 1 x 10-10 M) and the time of exposure to mifepristone (12 to 72 hours) to optimize or modulate expression for your cell line.
Introduction
Once you have established that your construct can be inducibly expressed, you may create a stable cell line that inducibly expresses your gene of interest. The pGene/V5-His and the pSwitch vectors contain the Zeocin™ resistance gene and the hygromycin resistance gene, respectively, to allow selection of stable cell lines using the selection agents, Zeocin™ and hygromycin B. We recommend that you first generate a stable cell line that expresses only the GAL4-DBD/hPR-LBD/p65-AD regulatory fusion protein, then use that cell line to create a second cell line which will express your gene of interest from the inducible expression plasmid. (see Note below). Alternatively, you can cotransfect with both plasmids (pSwitch and pGene/V5-His) and dual-select with hygromycin B and Zeocin™ to isolate a single stable cell line expressing both the regulatory fusion protein and your gene of interest.
The following section provides guidelines and instructions to generate stable cell lines by transfection.
Your gene of interest should not be expressed if you transfect your pGene/V5-His construct into mammalian host cells prior to transfecting the pSwitch plasmid. Mammalian cells do not contain endogenous GAL4 protein; therefore, transcription of the gene of interest should not be induced unless the GAL4-DBD/hPR-LBD/p65-AD regulatory fusion protein is present. While the E1b TATA sequence may not be completely silent, the basal transcription from the pGene/V5-His plasmid is generally undetectable.
Reminder: When generating a stable cell line expressing the GAL4-DBD/hPR-LBD/p65-AD regulatory fusion protein from pSwitch, select for clones that express the lowest levels of the GeneSwitch™ protein in the absence of mifepristone and the highest levels after induction with mifepristone. You may screen stable pSwitch clones for low basal expression and high inducibility by transient transfection with the pGene/V5-His/lacZ control plasmid.
Zeocin™
The pGene/V5-His plasmid contains the Zeocin™ resistance gene to allow selection of stable transfectants using Zeocin™.
Hygromycin B
The pSwitch vector contains the E. coli hygromycin resistance gene (HPH) (Gritz and Davies, 1983) for selection of transfectants using hygromycin B (Palmer et al., 1987). When added to cultured mammalian cells, hygromycin B acts as an aminocyclitol to inhibit protein synthesis by disrupting translocation and promoting mistranslation. Hygromycin B is supplied with the GeneSwitch™ Complete kit and is also available separately.
Hygromycin B is light sensitive. Store the liquid stock solution at +4°C protected from exposure to light.
Hygromycin is toxic. Do not ingest solutions containing the drug. Wear gloves, a laboratory coat, and safety glasses or goggles when handling hygromycin B and hygromycin B-containing solutions.
Preparing and Storing Hygromycin B
The hygromycin B included with the GeneSwitch™ Complete kit is supplied as a 100 mg/ml stock solution in autoclaved, deionized water and is filter-sterilized. The solution is brown in color. The stability of hygromycin B is guaranteed for six months, if stored at +4°C. Medium containing hygromycin B is stable for up to six weeks.
Determining Antibiotic Sensitivity
To successfully generate a stable cell line expressing the GAL4-DBD/hPR-LBD/p65-AD regulatory protein and your recombinant protein of interest, you need to determine the minimum concentration of each selection agent (Zeocin™ and hygromycin B) required to kill your untransfected host cell line. For each selection agent, test a range of concentrations (see below) to ensure that you determine the minimum concentration necessary for your cell line. Use the protocol below to determine the minimal concentrations of Zeocin™ and hygromycin B required to kill the parental cell line. See above for instructions to prepare and store hygromycin B.
Note: Typically, concentrations ranging from 50 to 1000 mg/ml Zeocin™ and 10 to 400 mg/ml hygromycin B are sufficient to kill most untransfected mammalian cell lines.
- Plate or split a confluent plate so the cells will be approximately 25% confluent. For each selection agent, prepare a set of 7 plates. Allow cells to adhere overnight.
- The next day, add the following concentrations of antibiotic to each plate in a set:
- For Zeocin™ selection, test 0, 50, 125, 250, 500, 750, and 1000 mg/ml Zeocin™
- For hygromycin selection, test 0, 10, 50, 100, 200, 400, 600 mg/ml hygromycin B
- Replenish the selective media every 3-4 and observe the percentage of surviving cells.
- Note the percentage of surviving cells at regular intervals to determine the appropriate concentration of each selection agent that kills the cells within 1-2 weeks after addition of the antibiotic.
Effect of Zeocin™ on Sensitive and Resistant Cells
Zeocin™'s method of killing is quite different from other antibiotics including hygromycin B, Geneticin®, and blasticidin. Cells do not round up and detach from the plate. Sensitive cells may exhibit the following morphological changes upon exposure to Zeocin™:
- Vast increase in size (similar to the effects of cytomegalovirus infecting permissive cells) and abnormal cell shape
- Presence of large empty vesicles in the cytoplasm (breakdown of the endoplasmic reticulum and Golgi apparatus, or other scaffolding proteins)
- Breakdown of plasma and nuclear membrane (appearance of many holes in these membranes)
Eventually, these "cells" will completely break down and only "strings" of protein remain. Zeocin™-resistant cells should continue to divide at regular intervals to form distinct colonies. There should not be any distinct morphological changes in Zeocin™-resistant cells when compared to cells not under selection with Zeocin™.
Possible Sites to Linearize pSwitch
To obtain stable transfectants, we recommend that you linearize the pSwitch plasmid before transfection. While linearizing the vector may not improve the efficiency of transfection, it increases the chances that the vector does not integrate in a way that disrupts the GAL4-DBD/hPR-LBD/p65-AD gene fusion or other elements necessary for expression in mammalian cells. The table below lists unique sites that may be used to linearize the pSwitch plasmid prior to transfection. Other restriction sites are possible.
Enzyme | Restriction Site (bp) | Location | Supplier |
Bst1107 I
|
5128
|
End of SV40 polyA
|
AGS
*, Fermentas, Takara, Roche Molecular Biochemicals
|
Sap I
|
5391
|
Backbone
|
New England Biolabs
|
BspLU11 I
|
5507
|
Backbone
|
Roche Molecular Biochemicals
|
Fsp I
|
6622
|
Ampicillin gene
|
Many
|
Possible Sites to Linearize pGene/V5-His A, B, and C
We recommend that you also linearize your pGene/V5-His construct before transfection. The table below lists unique sites that may be used to linearize the plasmid prior to transfection. Other restriction sites are possible. Note that the cleavage site is indicated for versions A, B, and C of pGene/V5-His. Be sure that your insert does not contain the restriction enzyme site you wish to use to linearize your vector.
Enzyme | Restriction Site (bp) (A,B,C) | Location | Supplier |
Bst1107 I
|
2439, 2443, 2435
|
End of SV40 polyA
|
AGS
*, Fermentas, Takara, Roche Molecular Biochemicals
|
Sap I
|
2702, 2706, 2698
|
Backbone
|
New England Biolabs
|
BspLU11 I
|
2818, 2822, 2814
|
Backbone
|
Roche Molecular Biochemicals
|
Eam1105 I
|
3708, 3712, 3704
|
Ampicillin gene
|
AGS
*, Fermentas, Takara
|
Fsp I
|
3930, 3934, 3926
|
Ampicillin gene
|
Many
|
Pvu I
|
4078, 4082, 4074
|
Ampicillin gene
|
Many
|
Ssp I
|
4512, 4516, 4508
| bla promoter
|
Many
|
*Angewandte Gentechnologie Systeme
Selecting Stable Integrants
Once you have determined the appropriate Zeocin™ and hygromycin B concentrations to use for selection, you can generate a stable cell line expressing pSwitch and your pGene/V5-His construct. We recommend that you first generate a stable cell line expressing pSwitch, and then use this cell line as the host for your pGene/V5-His construct. If you wish to perform cotransfection and dual selection of stable integrants, see below.
- Transfect the mammalian cell line of choice with pSwitch using the method of choice. Include a plate of untransfected cells as a negative control.
- 24 hours after transfection, wash the cells and add fresh medium to the cells.
- 48 hours after transfection, split the cells into fresh medium. Split the cells such that they are no more than 25% confluent. If the cells are too dense, the antibiotic will not kill the cells.
- Incubate the cells at 37°C for at least 2-3 hours until they have attached to the culture dish. Remove the medium and add fresh medium containing hygromycin B at the pre-determined concentration required for your cell line.
- Feed the cells with selective medium every 3-4 days until foci can be identified.
- Pick at least 20 hygromycin-resistant foci and expand each clone to test for mifepristone-inducible gene expression by transiently transfecting with the pGene/V5-His/lacZ positive control plasmid. Screen for those clones which exhibit the lowest levels of basal transcription and the highest levels of beta-galactosidase expression after addition of mifepristone.
- Once you have obtained a stable cell line expressing the GAL4-DBD/hPR-LBD/p65-AD fusion protein, you can use this cell line to isolate a stable cell line expressing your gene of interest from the pGene/V5-His construct. Repeat Steps 1 through 6, above, using your pGene/V5-His construct and Zeocin™ to select foci. Remember to maintain your cells in medium containing hygromycin as well.
- Pick and expand at least 20 foci to test for mifepristone-regulated gene expression.
Dual Selection of Stable Integrants
We recommend using the protocol above to generate double stable cell lines. However, if you wish to perform dual selection, you may cotransfect both pSwitch and your pGene/V5-His expression plasmid into the cell line of choice at a 1:1 ratio, and select for double stable transfectants using hygromycin B and Zeocin™ . (see Note below). Screen at least 40 foci for mifepristone-regulated expression of your gene of interest using an appropriate assay for your protein (e.g. western blot, enzymatic assay).
Some cells may be more sensitive to dual selection than selection with a single antibiotic; therefore, you may find that you can use lower concentrations of hygromycin B and Zeocin™ for dual selection of transfectants. Once you have isolated stable cell lines containing both pSwitch and your pGene/V5-His construct and have tested for mifepristone-inducible expression of your gene of interest, we recommend that you perform a time course of mifepristone induction to optimize expression of your protein of interest (e.g. 0, 12, 24, 48, 72 hours, etc.). Use the appropriate concentration of mifepristone for your cell line as previously determined. We have observed as much as 50-200-fold induction of b-galactosidase in a double stable cell line (NIH3T3 cells transfected with pSwitch and pGene/V5-His/lacZ) after 24 hours of treatment with 1 x 10-8 M mifepristone. Note that your induction levels may vary depending on the nature of your gene of interest and the particular clone that you choose.
After 24 hours of induction with 1 x 10-8 M mifepristone, we have observed continuous expression of b-galactosidase in a double stable cell line (NIH3T3 cells transfected with pSwitch and pGene/V5-His/lacZ) even after removal of mifepristone from the tissue culture medium. High levels of beta-galactosidase protein are still detectable after culturing cells for 144 hours in the absence of mifepristone. If you wish to perform repetitive induction experiments with your gene of interest, the GeneSwitch™ System may not be appropriate for this use. Empirical experimentation will be necessary as results may vary depending on the nature of the gene of interest and the host cell line.
Preparing Cells for Lysis
Use the procedure below to prepare cells for lysis prior to purification of your protein on ProBond™ or Ni-NTA. You will need approximately 5 x 106 to 1 x 107 stably transfected cells to purify your protein on a 2 ml ProBond™ or Ni-NTA column.
- Seed cells in either five T-75 flasks or 2 to 3 T-175 flasks.
- Grow the cells in selective medium until they are approximately 50% confluent.
- Add the appropriate concentration of mifepristone and induce expression of your protein of interest to the desired level.
- Harvest the cells by treating with trypsin-EDTA for 2 to 5 minutes or by scraping the cells in PBS.
- Inactivate the trypsin by diluting with fresh medium (if necessary) and transfer the cells to a sterile microcentrifuge tube.
- Centrifuge the cells at 1500 rpm for 5 minutes. Resuspend the cell pellet in PBS.
- Centrifuge the cells at 1500 rpm for 5 minutes. You may lyse the cells immediately or freeze in liquid nitrogen and store at –70°C until needed. Refer to the ProBond™ or Ni-NTA manual for guidelines about sample preparation
The pGene/V5-His A, B, and C vectors allow cloning of your gene of interest in frame with a C-terminal peptide. Expression of the gene of interest is controlled by a GAL4 UAS and an E1b TATA box. The vector also contains a synthetic intron to enhance expression of your gene. The complete nucleotide sequences for pGene/V5-His A, B, and C are available for download on our website or from Technical Service.
Features of pGene/V5-His
The table below describes the relevant features of pGene/V5-His. All features have been functionally tested.
| Feature | Benefit |
| GAL4 Upstream Activating Sequences (UAS) | Contains six copies of a 17 bp sequence that allows binding and transcriptional activation of the gene of interest by the GAL4 DBD/hPR-LBD/p65 AD fusion protein (Giniger et al., 1985; Wang et al., 1994) |
| Adenovirus E1b TATA sequence | Permits transcriptional initiation of the gene of interest (Lillie and Green, 1989) |
| Synthetic intron IVS8 | Enhances expression of the gene of interest |
| pGene forward priming site | Allows sequencing in the sense orientation |
| Multiple cloning site | Allows insertion of your gene of interest |
| V5 epitope | (Gly-Lys-Pro-Ile-Pro-Asn-Pro-Leu-Leu-Gly-Leu-Asp-Ser-Thr) Allows detection of your recombinant protein with the Anti-V5 antibodies (Southern et al., 1991) |
| C-terminal polyhistidine (6xHis) tag | Permits purification of your recombinant protein on metal-chelating resin such as ProBond™ or Ni-NTA. In addition, the C-terminal polyhistidine tag is the epitope for the Anti-His(C-term) antibodies (Lindner et al., 1997) |
| Bovine growth hormone (BGH) polyadenylation sequence | Permits efficient polyadenylation of mRNA (Goodwin and Rottman, 1992) |
| BGH reverse priming site | Permits sequencing of the non-coding strand |
| f1 origin | Allows rescue of single-stranded DNA |
| SV40 early promoter and origin | Allows efficient, high-level expression of the Zeocin™ resistance gene in mammalian cells and episomal replication in cells expressing SV40 large T antigen |
| EM7 promoter | Synthetic prokaryotic promoter for expression of the Zeocin™ resistance gene in E. coli |
| Zeocin™ resistance (Sh ble) gene | Permits selection of stable transfectants in mammalian cells (Mulsant et al., 1988) and transformants in E. coli (Drocourt et al., 1990) |
| SV40 early polyadenylation signal | Allows polyadenylation of mRNA |
| pUC origin | Permits high-copy number replication and maintenance in E. coli |
| bla promoter | Allows expression of the ampicillin (bla) resistance gene |
Ampicillin (
bla) resistance gene
(ß-lactamase)
| Allows selection of transformants in E. coli |
Map of pSwitch is a 7323 bp vector that expresses a 73 kDa fusion protein consisting of the yeast GAL4 DNA binding domain (DBD), a truncated human progesterone receptor ligand binding domain (hPR-LBD), and the human NF-kB p65 activation domain (AD) under the control of GAL4 upstream activating sequences (UAS) and a Herpes Simplex Virus thymidine kinase (TK) minimal promoter. The vector also includes a synthetic intron to enhance expression of the fusion gene. The complete sequence for pSwitch is available for download or by contacting Technical Service.
Features of pSwitch
The table below describes the relevant features of pSwitch. All features have been functionally tested and the vector has been fully sequenced.
| Feature | Benefit |
| GAL4 upstream activating sequences (UAS) |
Contains four copies of a 17 bp sequence that allows binding and transcriptional activation of the gene of interest by the GAL4-DBD/hPR-LBD/p65-AD fusion protein (Giniger
et al., 1985; Wang
et al., 1994)
|
| Herpes Simplex Virus thymidine kinase (TK) minimal promoter |
Allows expression of the
GAL4-DBD/hPR-LBD/p65-AD fusion gene (McKnight, 1980)
|
| Synthetic intron IVS8 |
Enhances expression of the
GAL4-DBD/hPR-LBD/p65-AD fusion gene
|
| GAL4-DBD/hPR-LBD/p65-AD fusion gene |
Encodes a 73 kDa fusion protein containing the yeast GAL4 DNA binding domain (Laughon and Gesteland, 1984; Marmorstein
et al., 1992), a truncated human progesterone receptor ligand binding domain (Kastner
et al., 1990; Misrahi
et al., 1987; Wang
et al., 1994), and the human NF-kB p65 activation domain (Burcin
et al., 1999; Deloukas and Loon, 1993; Ruben
et al., 1991) to allow mifepristone-regulated expression of the gene of interest
|
| Bovine growth hormone (BGH) polyadenylation sequence |
Permits efficient polyadenylation of mRNA (Goodwin and Rottman, 1992)
|
| BGH reverse priming site |
Permits sequencing of the non-coding strand
|
| f1 origin |
Allows rescue of single-stranded DNA
|
| SV40 early promoter and origin |
Allows efficient, high-level expression of the hygromycin resistance gene in mammalian cells and episomal replication in cells expressing SV40 large T antigen
|
| Hygromycin (HPH) resistance gene |
Permits selection of stable transfectants in mammalian cells (Gritz and Davies, 1983)
|
| SV40 early polyadenylation signal |
Allows polyadenylation of mRNA
|
| pUC origin |
Permits high-copy number replication and maintenance in
E. coli |
| bla promoter |
Allows expression of the ampicillin (
bla) resistance gene
|
Ampicillin (
bla) resistance gene
(ß-lactamase)
|
Allows selection of transformants in
E. coli |
GAL4 DNA Binding Domain
The Saccharomyces cerevisiae GAL4 gene encodes a transcription factor that activates expression of genes required for galactose catabolism in yeast (Laughon and Gesteland, 1984). The GAL4 protein regulates transcription of target genes by binding as a homodimer to specific 17 nucleotide GAL4 binding sites contained within the upstream activating sequences (UAS) of these genes (Giniger et al., 1985; Marmorstein et al., 1992). The DNA binding function of GAL4 has been mapped to the N-terminal portion of the protein (Carey et al., 1989).
The GAL4-DBD/hPR-LBD/p65-AD fusion gene contains a DNA fragment encoding amino acids 1-93 of the GAL4 gene. This portion of the GAL4 protein includes regions that have been shown to be responsible for DNA binding (amino acids 1-65) (Keegan et al., 1986), dimerization (amino acids 65-93) (Carey et al., 1989), and nuclear localization (amino acids 1-29) (Nelson and Silver, 1989).
Human Progesterone Receptor Ligand-Binding Domain
The human progesterone receptor (Kastner et al., 1990; Misrahi et al., 1987) is a member of the steroid and thyroid hormone receptor superfamily (Evans, 1988; Truss and Beato, 1993). In the absence of ligand, the progesterone receptor is localized in the nucleus in an inactive form (Guiochon-Mantel et al., 1989; Perrot-Applanat et al., 1985). In the presence of its cognate ligand, progesterone and other progesterone agonists, the progesterone receptor undergoes a conformational change to an active form. The ligand-bound receptor then homodimerizes and functions as a transcription factor to regulate expression of genes involved in cellular proliferation and differentiation (Evans, 1988; Simons, 1998). The ligand-binding functions of the progesterone receptor reside in the C-terminal portion of the protein (amino acids 640-933) (Vegeto et al., 1992). Synthetic progesterone antagonists including mifepristone also bind to the progesterone receptor in this region (Vegeto et al., 1992).
The portion of the ligand-binding domain included in the GAL4-DBD/hPR-LBD/p65-AD fusion gene encodes amino acids 640-914 of the human progesterone receptor. This hPR-LBD fragment contains a deletion of 19 amino acids from the C-terminal end of the native hPR-LBD, which allows the truncated hPR-LBD to bind with high affinity to mifepristone and other synthetic progesterone antagonists, but abolishes its ability to bind to progesterone or other endogenous steroid hormones (Wang et al., 1994; Wang et al., 1997).
Human p65 Activation Domain
The human p65 protein, originally identified as a relative of the rel oncogene, encodes a subunit of the second messenger, NF-kB (Ruben et al., 1991). NF-kB consists of a heterodimer between two proteins, p65 and p50, and functions as a pleiotrophic transcriptional activator in eukaryotes (Baeuerle, 1991). In its inactive form, NF-kB is localized in the cytoplasm as a complex with the inhibitor, IkB (Baeuerle and Baltimore, 1988). Binding of IkB to NF-kB occurs via the p65 protein. In addition to its interaction with IkB, the p65 protein is also responsible for the transcription activation function of NF-kB (Schmitz and Baeuerle, 1991). The portion of p65 included in the GAL4-DBD/hPR-LBD/p65-AD fusion gene encodes amino acids 283-551 of the protein. Two distinct transactivation domains have been identified within this region of the protein (Schmitz and Baeuerle, 1991).
Mifepristone (11b-[4-dimethylamino]phenyl-17b-hydroxy-17-[1-propynyl]estra-4,9-dien-3-one) is a synthetic 19-norsteroid that binds with high affinity to the human progesterone receptor (Kd <1 x 10-9 M) and the glucocorticoid receptor (Kd <1 x 10-9 M) and acts as both a progesterone and glucocorticoid antagonist (Baulieu, 1989). Mifepristone is also known as RU 486 and has been used as an abortifacient at high doses (3-10 mg/kg) (Baulieu, 1989; Philibert et al., 1985).
In the GeneSwitch™ System, mifepristone acts as an agonist to activate gene transcription by binding to a truncated human progesterone receptor (Wang et al., 1997). The concentrations of mifepristone used to induce gene expression in the GeneSwitch™ System are extremely low (1 x 10-8 M), non-toxic, and exert no known pleiotrophic effects on mammalian cells that lack endogenous progesterone and glucocorticoid receptors (Wang et al., 1994).
Molecular Weight, Formula, and Structure
The formula for mifepristone is C29H35NO2 and the molecular weight is 429.6. The diagram below shows the structure of mifepristone.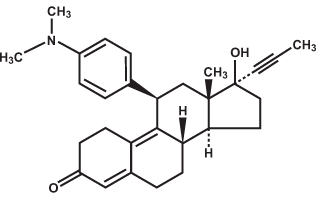
Handling Mifepristone
- Mifepristone is toxic. Do not ingest or inhale the powder or solutions containing the drug.
- Use caution when handling large quantities of mifepristone (> 100 mg). At high doses (see above), mifepristone may impair fertility and may cause harm to the unborn child.
- Always wear gloves, a laboratory coat, and safety glasses or goggles when handling mifepristone and mifepristone-containing solutions.
Preparing and Storing Stock Solutions
Mifepristone is supplied with the GeneSwitch™ Complete and Core Systems, but may also be obtained separately (Catalog no. H110-01) in 100 mg aliquots. To prepare mifepristone:
- Resuspend the 100 mg aliquot of mifepristone in 233 ml of 100% ethanol to make a 1 mM (1 x 10-3 M) stock solution of mifepristone that is clear. Do not heat.
- Dilute 100 ml of the 1 mM mifepristone stock solution in 10 ml of 100% ethanol to make a 10 mM (1 x 10-5 M) working solution of mifepristone. Use this working solution when adding mifepristone to your cells.
- Store the 1 mM stock solution and the 10 mM working solution of mifepristone at -20°C to prevent evaporation. The solutions are stable indefinitely.
Zeocin™ is a member of the bleomycin/phleomycin family of antibiotics isolated from Streptomyces. Antibiotics in this family are broad spectrum antibiotics that act as strong anti-bacterial and anti-tumor drugs. They show strong toxicity against bacteria, fungi (including yeast), plants, and mammalian cells (Baron et al., 1992; Drocourt et al., 1990; Mulsant et al., 1988; Perez et al., 1989).
The Zeocin™ resistance protein has been isolated and characterized (Calmels et al., 1991; Drocourt et al., 1990). This protein, the product of the Sh ble gene (Streptoalloteichus hindustanus bleomycin gene), is a 13.7 kDa protein that binds Zeocin™ and inhibits its DNA strand cleavage activity. Expression of this protein in eukaryotic and prokaryotic hosts confers resistance to Zeocin™.
Molecular Weight, Formula, and Structure
The formula for Zeocin™ is C60H89N21O21S3 and the molecular weight is 1,535. The diagram below shows the structure of Zeocin™.

Applications of Zeocin™
Zeocin™ is used for selection in mammalian cells (Mulsant et al., 1988); plants (Perez et al., 1989); yeast (Baron et al., 1992); and prokaryotes (Drocourt et al., 1990). Suggested concentrations of Zeocin™ for selection in mammalian cell lines and E. coli are listed below:
| Organism | Zeocin™ Concentration and Selective Medium |
| E. coli | 25-50 µg/ml in Low Salt LB medium* |
| Mammalian Cells | 50-1000 µg/ml (varies with cell line) |
*Efficient selection requires that the concentration of NaCl be no more than 5 g/L (< 90 mM)
Handling Zeocin™
- High salt and acidity or basicity inactivate Zeocin™ therefore, we recommend that you reduce the salt in bacterial medium and adjust the pH to 7.5 to keep the drug active (see Low Salt LB Medium). Note that the salt concentration and pH do not need to be adjusted when preparing tissue culture medium containing Zeocin™.
- Store Zeocin™ at -20°C and thaw on ice before use.
- Zeocin™ is light sensitive. Store drug, plates, and medium containing drug in the dark.
- Wear gloves, a laboratory coat, and safety glasses or goggles when handling solutions containing Zeocin™.
- Zeocin™ is toxic. Do not ingest or inhale solutions containing the drug.
- Store tissue culture medium containing Zeocin™ at +4°C in the dark. Medium containing Zeocin™ is stable for 1-2 months.
Ordering Information
Zeocin™ can be purchased from Thermo Fisher Scientific. For your convenience, the drug is prepared in autoclaved, deionized water and available in 1.25 ml aliquots at a concentration of 100 mg/ml. The Zeocin™ solution is blue in color. The stability of Zeocin™ is guaranteed for six months, if stored at -20°C.
Amount Catalog no.
1 gram R250-01
5 grams R250-05
LB (Luria-Bertani) Medium and Plates Composition:
10 g Tryptone
10 g NaCl
5 g Yeast Extract
pH 7.0
1. Combine the reagents above and add deionized, distilled water to 950 ml.
2. Adjust the pH of the solution to 7.0 with NaOH and bring the volume up to 1 liter.
3. Autoclave on liquid cycle for 20 minutes at 15 psi. Allow solution to cool to 55°C and add antibiotic if needed.
4. Store at room temperature or at +4°C.
LB agar plates
1. Prepare LB medium as above, but add 15 g/L agar before autoclaving.
2. Autoclave on liquid cycle for 20 minutes at 15 psi.
3. After autoclaving, cool to ~55°C, add antibiotic (i.e. 50-100 µg/ml of ampicillin), and pour into 10 cm plates.
4. Let harden, then invert and store at +4°C, in the dark.
L ow Salt LB Medium with Zeocin™
10 g Tryptone
5 g NaCl
5 g Yeast Extract
pH 7.5
1. Combine the reagents above and add deionized, distilled water to 950 ml.
2. Adjust pH to 7.5 with 5 M NaOH. Bring the volume up to 1 liter. For plates, add 15 g/L agar before autoclaving.
3. Autoclave on liquid cycle at 15 lbs/sq. in. and 121°C for 20 minutes.
4. Thaw Zeocin™ on ice and vortex before removing an aliquot.
5. Allow the medium to cool to at least 55°C before adding Zeocin™ to a 25 mg/ml final concentration.
6. Store plates at +4°C in the dark. Plates containing Zeocin™ are stable for 1-2 weeks.
Cell Lysis Buffer
50 mM Tris, pH 7.8
150 mM NaCl
1% Nonidet P-40
1. This solution can be prepared from the following common stock solutions.
For 100 ml, combine:
1 M Tris base 5 ml
5 M NaCl 3 ml
Nonidet P-40 1 ml
2. Bring the volume up to 90 ml with deionized water and adjust the pH to 7.8 with HCl.
3. Bring the volume up to 100 ml. Store at room temperature.
Note: Protease inhibitors may be added at the following concentrations:
1 mM PMSF
1 mg/ml pepstatin
1 mg/ml leupeptin
10 g Tryptone
10 g NaCl
5 g Yeast Extract
pH 7.0
1. Combine the reagents above and add deionized, distilled water to 950 ml.
2. Adjust the pH of the solution to 7.0 with NaOH and bring the volume up to 1 liter.
3. Autoclave on liquid cycle for 20 minutes at 15 psi. Allow solution to cool to 55°C and add antibiotic if needed.
4. Store at room temperature or at +4°C.
LB agar plates
1. Prepare LB medium as above, but add 15 g/L agar before autoclaving.
2. Autoclave on liquid cycle for 20 minutes at 15 psi.
3. After autoclaving, cool to ~55°C, add antibiotic (i.e. 50-100 µg/ml of ampicillin), and pour into 10 cm plates.
4. Let harden, then invert and store at +4°C, in the dark.
L ow Salt LB Medium with Zeocin™
10 g Tryptone
5 g NaCl
5 g Yeast Extract
pH 7.5
1. Combine the reagents above and add deionized, distilled water to 950 ml.
2. Adjust pH to 7.5 with 5 M NaOH. Bring the volume up to 1 liter. For plates, add 15 g/L agar before autoclaving.
3. Autoclave on liquid cycle at 15 lbs/sq. in. and 121°C for 20 minutes.
4. Thaw Zeocin™ on ice and vortex before removing an aliquot.
5. Allow the medium to cool to at least 55°C before adding Zeocin™ to a 25 mg/ml final concentration.
6. Store plates at +4°C in the dark. Plates containing Zeocin™ are stable for 1-2 weeks.
Cell Lysis Buffer
50 mM Tris, pH 7.8
150 mM NaCl
1% Nonidet P-40
1. This solution can be prepared from the following common stock solutions.
For 100 ml, combine:
1 M Tris base 5 ml
5 M NaCl 3 ml
Nonidet P-40 1 ml
2. Bring the volume up to 90 ml with deionized water and adjust the pH to 7.8 with HCl.
3. Bring the volume up to 100 ml. Store at room temperature.
Note: Protease inhibitors may be added at the following concentrations:
1 mM PMSF
1 mg/ml pepstatin
1 mg/ml leupeptin
- Abruzzese, R. V., Godin, D., Burcin, M., Mehta, V., French, M., Li, Y., O'Malley, B. W., and Nordstrom, J. L. (1999). Ligand-Dependent Regulation of Plasmid-Based Transgene Expression in Vivo. Human Gene Therapy 10, 1499-1507.
- Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Smith, J. A., and Struhl, K. (1994). Current Protocols in Molecular Biology (New York: Greene Publishing Associates and Wiley-Interscience).
- Baeuerle, P. A. (1991). The Inducible Transcription Activator NF-kappa B: Regulation by Distinct Protein Subunits. Biochim. Biophys. Acta 1072, 63-80.
- Baeuerle, P. A., and Baltimore, D. (1988). I Kappa B: A Specific Inhibitor of the NF-kappa B Transcription Factor. Science 242, 540-546.
- Baron, M., Reynes, J. P., Stassi, D., and Tiraby, G. (1992). A Selectable Bifunctional b-Galactosidase: Phleomycin-resistance Fusion Protein as a Potential Marker for Eukaryotic Cells. Gene 114, 239-243.
- Baulieu, E. E. (1989). Contragestion and Other Clinical Applications of RU486, an Antiprogesterone at the Receptor. Science 245, 1351-1357.
- Burcin, M. M., Schiedner, G., Kochanek, S., Tsai, S. Y., and O'Malley, B. W. (1999). Adenovirus-Mediated Regulable Target Gene Expression in vivo. Proc. Natl. Acad. Sci. USA 96, 355-360.
- Calmels, T., Parriche, M., Burand, H., and Tiraby, G. (1991). High Efficiency Transformation of Tolypocladium geodes Conidiospores to Phleomycin Resistance. Curr. Genet. 20, 309-314.
- Carey, M., Kakidani, H., Leatherwood, J., Mostashari, F., and Ptashne, M. (1989). An Amino-Terminal Fragment of GAL4 Binds DNA as a Dimer. J. Mol. Biol. 209, 423-432.
- Chen, C., and Okayama, H. (1987). High-Efficiency Transformation of Mammalian Cells by Plasmid DNA. Mol. Cell. Biol. 7, 2745-2752.
- Chu, G., Hayakawa, H., and Berg, P. (1987). Electroporation for the Efficient Transfection of Mammalian Cells with DNA. Nucleic Acids Res. 15, 1311-1326.
- DeFranco, D. B. (1998) Subcellular and Subnuclear Trafficking of Steroid Receptors. In Molecular Biology of Steroid and Nuclear Hormone Receptors, L. P. Freedman, ed. (Boston, MA: Birkhauser), pp. 19-34.
- Deloukas, P., and Loon, A. P. V. (1993). Genomic Organization of the Gene Encoding the p65 Subunit of NF-kappa B: Multiple Variants of the p65 Protein May be Generated by Alternative Splicing. Hum. Mol. Genet. 2, 1895-1900.
- Drocourt, D., Calmels, T. P. G., Reynes, J. P., Baron, M., and Tiraby, G. (1990). Cassettes of the Streptoalloteichus hindustanus ble Gene for Transformation of Lower and Higher Eukaryotes to Phleomycin Resistance. Nucleic Acids Res. 18, 4009.
- Evans, R. M. (1988). The Steroid and Thyroid Hormone Receptor Superfamily. Science 240, 889-895.
- Felgner, P. L., Holm, M., and Chan, H. (1989). Cationic Liposome Mediated Transfection. Proc. West. Pharmacol. Soc. 32, 115-121.
- Felgner, P. L. a., and Ringold, G. M. (1989). Cationic Liposome-Mediated Transfection. Nature 337, 387-388.
- Gasc, J. M., Delahaye, F., and Baulieu, E. E. (1989). Compared Intracellular Localization of the Glucocorticosteroid and Progesterone Receptors: An Immunocytochemical Study. Exp. Cell Res. 181, 492-504.
- Giniger, E., Varnum, S. M., and Ptashne, M. (1985). Specific DNA Binding of GAL4, a Positive Regulatory Protein of Yeast. Cell 40, 767-774
- Goodwin, E. C., and Rottman, F. M. (1992). The 3´-Flanking Sequence of the Bovine Growth Hormone Gene Contains Novel Elements Required for Efficient and Accurate Polyadenylation. J. Biol. Chem. 267, 16330-16334.
- Gritz, L., and Davies, J. (1983). Plasmid-Encoded Hygromycin-B Resistance: The Sequence of Hygromycin-B-Phosphotransferase Gene and its Expression in E. coli and S. cerevisiae. Gene 25, 179-188.
- Guiochon-Mantel, A., Loosfelt, H., Lescop, P., Sar, S., Atger, M., Perrot-Applanat, M., and Milgrom, E. (1989). Mechanisms of Nuclear Localization of the Progesterone Receptor: Evidence for Interaction between Monomers. Cell 57, 1147-1154.
- Kastner, P., Krust, A., Turcotte, B., Stropp, U., Tora, L., Gronemeyer, H., and Chambon, P. (1990). Two Distinct Estrogen-Regulated Promoters Generate Transcripts Encoding the Two Functionally Different Human Progesterone Receptor Forms A and B. EMBO J. 9, 1603-1614.
- Keegan, L., Gill, G., and Ptashne, M. (1986). Separation of DNA Binding from the Transcription-Activating Function of a Eukaryotic Regulatory Protein. Science 231, 699-704.
- Kozak, M. (1987). An Analysis of 5´-Noncoding Sequences from 699 Vertebrate Messenger RNAs. Nucleic Acids Res. 15, 8125-8148. v
- Kozak, M. (1991). An Analysis of Vertebrate mRNA Sequences: Intimations of Translational Control. J. Cell Biology 115, 887-903.
- Kozak, M. (1990). Downstream Secondary Structure Facilitates Recognition of Initiator Codons by Eukaryotic Ribosomes. Proc. Natl. Acad. Sci. USA 87, 8301-8305.
- Laughon, A., and Gesteland, R. F. (1984). Primary Structure of the Saccharomyces cerevisiae GAL4 Gene. Mol. Cell. Biol. 4, 260-267.
- Lillie, J. W., and Green, M. R. (1989). Transcription Activation by the Adenovirus E1a Protein. Nature 338, 39-44.
- Lindner, P., Bauer, K., Krebber, A., Nieba, L., Kremmer, E., Krebber, C., Honegger, A., Klinger, B., Mocikat, R., and Pluckthun, A. (1997). Specific Detection of His-tagged Proteins With Recombinant Anti-His Tag scFv-Phosphatase or scFv-Phage Fusions. BioTechniques 22, 140-149.
- Marmorstein, R., Carey, M., Ptashne, M., and Harrison, S. C. (1992). DNA Recognition by GAL4: Structure of a Protein-DNA Complex. Nature 356, 408-414.
- McKnight, S. L. (1980). The Nucleotide Sequence and Transcript Map of the Herpes Simplex Virus Thymidine Kinase Gene. Nucleic Acids Res. 8, 5949-5964.
- Miller, J. H. (1972). Experiments in Molecular Genetics (Cold Spring Harbor, New York: Cold Spring Harbor Laboratory).
- Misrahi, M., Atger, M., d'Auriol, L., Loosfelt, H., Meriel, C., Fridlansky, F., Guiochon-Mantel, A., Galibert, F., and Milgrom, E. (1987). Complete Amino Acid Sequence of the Human Progesterone Receptor Deduced from Cloned cDNA. Biochem. Biophys. Res. Commun. 143, 740-748.
- Mulsant, P., Tiraby, G., Kallerhoff, J., and Perret, J. (1988). Phleomycin Resistance as a Dominant Selectable Marker in CHO Cells. Somat. Cell Mol. Genet. 14, 243-252.
- Nelson, M., and Silver, P. (1989). Context Affects Nuclear Protein Localization in Saccharomyces cerevisiae. Mol. Cell. Biol. 9, 384-389.
- Palmer, T. D., Hock, R. A., Osborne, W. R. A., and Miller, A. D. (1987). Efficient Retrovirus-Mediated Transfer and Expression of a Human Adenosine Deaminase Gene in Diploid Skin Fibroblasts from an Adenosine-Deficient Human. Proc. Natl. Acad. Sci. U.S.A. 84, 1055-1059.
- Perez, P., Tiraby, G., Kallerhoff, J., and Perret, J. (1989). Phleomycin Resistance as a Dominant Selectable Marker for Plant Cell Transformation. Plant Mol. Biol. 13, 365-373.
- Perrot-Applanat, M., Logeat, F., Groyer-Picard, M. T., and Milgrom, E. (1985). Immunocytochemical Study of Mammalian Progesterone Receptor Using Monoclonal Antibodies. Endocrinol. 116, 1473-1484.
- Philibert, D., Moguilewsky, M., Mary, I., Lecaque, D., Tournemine, C., Secchi, J., and Deraedt, R. (1985). In The Antiprogestin Steroid RU486 and Human Fertility Control, E. E. Baulieu and S. J. Segal, eds. (New York: Plenum), pp. 49-68.
- Ruben, S. M., Dillon, P. J., Schreck, R., Henkel, T., Chen, C. H., Maher, M., Baeuerle, P. A., and Rosen, C. A. (1991). Isolation of a Rel-related Human cDNA that Potentially Encodes the 65-kD Subunit of NF-kappa B. Science 251, 1490-1493.
- Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). Molecular Cloning: A Laboratory Manual, Second Edition (Plainview, New York: Cold Spring Harbor Laboratory Press).
- Schmitz, M. L., and Baeuerle, P. A. (1991). The p65 Subunit is Responsible for the Strong Transcription Activating Potential of NF-kappa B. EMBO J. 10, 3805-3817.
- Shigekawa, K., and Dower, W. J. (1988). Electroporation of Eukaryotes and Prokaryotes: A General Approach to the Introduction of Macromolecules into Cells. BioTechniques 6, 742-751.
- Simons, S. S., Jr. (1998) Structure and Function of the Steroid and Nuclear Receptor Ligand Binding Domain. In Molecular Biology of Steroid and Nuclear Hormone Receptors, L. P. Freedman, ed. (Boston, MA: Birkhauser), pp. 35-104.
- Southern, J. A., Young, D. F., Heaney, F., Baumgartner, W., and Randall, R. E. (1991). Identification of an Epitope on the P and V Proteins of Simian Virus 5 That Distinguishes Between Two Isolates with Different Biological Characteristics. J. Gen. Virol. 72, 1551-1557.
- Truss, M., and Beato, M. (1993). Steroid Hormone Receptors: Interaction with Deoxyribonucleic Acid and Transcription Factors. Endocr. Rev. 14, 459-479.
- Vegeto, E., Allan, G. F., Schrader, W. T., Tsai, M. J., McDonnell, D. P., and O'Malley, B. W. (1992). The Mechanism of RU486 Antagonism is Dependent on the Conformation of the Carboxy-Terminal Tail of the Human Progesterone Receptor. Cell 69, 703-713.
- Wang, Y., B.W. O'Malley, J., Tsai, S. Y., and O'Malley, B. W. (1994). A Regulatory System for Use in Gene Transfer. Proc. Natl. Acad. Sci. USA 91, 8180-8184.
- Wang, Y., Xu, J., Pierson, T., O'Malley, B. W., and Tsai, S. Y. (1997). Positive and Negative Regulation of Gene Expression in Eukaryotic Cells with an Inducible Transcriptional Regulator. Gene Therapy 4, 432-441.
- Wigler, M., Silverstein, S., Lee, L.-S., Pellicer, A., Cheng, Y.-C., and Axel, R. (1977). Transfer of Purified Herpes Virus Thymidine Kinase Gene to Cultured Mouse Cells. Cell 11, 223-232.
- Ylikomi, T., Bocquel, M. T., Berry, M., Gronemeyer, H., and Chambon, P. (1992). Cooperation of Proto-Signals for Nuclear Accumulation of Estrogen and Progesterone Receptors. EMBO J. 11, 3681-3694.
LT055