Search
Getting Started with RNAi In Vivo
RNA interference (RNAi) is a natural mechanism by which small interfering RNAs (siRNAs) mediate sequence-specific degradation of targeted RNA molecules. Technology exploiting this phenomenon may be used for functional genomics and target validation, and is being developed for therapeutic purposes. Because of its potency, siRNA has rapidly become the most widely used trigger for inducing gene knockdown in cultured mammalian cells. siRNA delivery in vivo is a more challenging task. This article describes an RNAi experiment in an animal model in which siRNA was administered using hydrodynamic tail vein injection.
Routes for Administering RNAi
A growing number of in vivo RNAi studies use mouse models. There are many reports of successful local administration of siRNA and antisense oligonucleotides. For example, several groups have focused on developing oligonucleotide-based therapeutics for eye-related disorders [1]. Because oligonucleotides are directly injected into the eye, the amount of material required is much smaller (and consequently less expensive) compared to that required for systemic drug delivery. Other advantages of targeting ocular tissue include lower RNase levels in the eye, compared to blood, and inherent host defense and clearance mechanisms that may promote cellular uptake of siRNA in the eye. Localized siRNA delivery to the respiratory system has also shown promise. Several research teams have demonstrated that intranasal administration of synthetic siRNA, with or without transfection agents, effectively treats influenza and respiratory syncytial virus [2, 3].
Problems Associated with Systemic Delivery
Since most target tissues or organs cannot be accessed by local administration of siRNA, the ultimate goal is a systemic delivery route by intravenous injection. Besides the practical considerations of delivering siRNA to an internal organ, inhibiting gene expression in multiple tissues (e.g., during treatment of highly metastatic tumors) is sometimes necessary. Success with systemic delivery has been limited by rapid blood clearance of molecules <10 nm in size, RNA degradation in the bloodstream, and poor cellular penetration by siRNA due to its large molecular weight and high negative charge.
Systemic siRNA Delivery to Liver
While additional work is required to develop effective technology for delivery of siRNA into many tissues through systemic administration, delivery to the liver of mice is rather straightforward. In this article, we describe siRNA delivery to the liver of mice by hydrodynamic (high-pressure) tail vein injection [4]. Because a large volume (equal to blood volume) is intravenously injected within a few seconds, contact time of siRNA with blood is limited, and the siRNA is almost quantitatively and specifically delivered to liver cells. In these experiments, siRNA and a reporter vector were co-injected, and knockdown of exogenous gene expression in tissue samples was measured at the mRNA and protein level [see sidebar, "Methods" and "Applications of the pMIR-REPORT™ miRNA Expression Reporter Vector System"].
When only the conrol pMIR-REPORT Luciferase Vector was injected, high levels of firefly luciferase (fLuc) expression were detected in liver samples, both at the protein and RNA levels. fLuc gene expression in kidneys, spleen, and lung was 100–1000X lower than in the liver, indicating targeted delivery of the reporter vector to the liver (data not shown). When the pMIR-REPORT Luciferase Vector, fLuc-specific Silencer® siRNA, and pMIR-REPORT b-galactosidase Vector (for normalization of delivery efficiency) were co-injected, significant reduction of fLuc protein (Figure 1A) and corresponding mRNA (Figure 1B) levels were observed. No reduction of fLuc was detected in the control group, which was injected with the nonspecific Silencer Negative Control #1 siRNA.
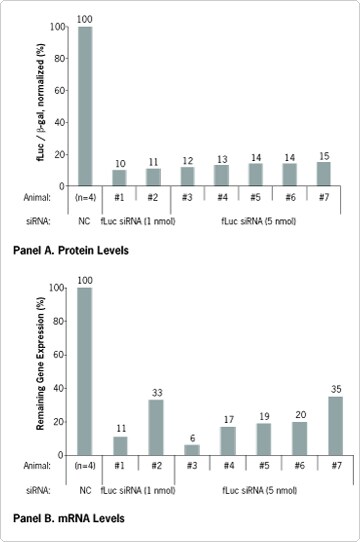
Figure 1. siRNA-induced Reduction of fLuc Protein and mRNA Levels in Mouse Liver. Ambion pMIR-REPORT™ Luciferase and β-galactosidase vectors and siRNA (1 or 5 nmol) that targets firefly luciferase (fLuc) were injected at constant high pressure into the tail veins of seven mice. Data were normalized to the average result from four mice injected with Silencer® Negative Control #1 siRNA. Panel A: fLuc and β-galactosidase (β-gal) protein levels were measured using the Tropix® Dual-Light® Luminescent Reporter Gene Assay System (Applied Biosystems). Each bar represents one animal, and readings were performed in triplicate. Panel B: TaqMan® Gene Expression Assays were used to measure fLuc mRNA levels. Each bar represents one animal, and readings were performed in triplicate and averaged.
The data demonstrated the following:
When only the conrol pMIR-REPORT Luciferase Vector was injected, high levels of firefly luciferase (fLuc) expression were detected in liver samples, both at the protein and RNA levels. fLuc gene expression in kidneys, spleen, and lung was 100–1000X lower than in the liver, indicating targeted delivery of the reporter vector to the liver (data not shown). When the pMIR-REPORT Luciferase Vector, fLuc-specific Silencer® siRNA, and pMIR-REPORT b-galactosidase Vector (for normalization of delivery efficiency) were co-injected, significant reduction of fLuc protein (Figure 1A) and corresponding mRNA (Figure 1B) levels were observed. No reduction of fLuc was detected in the control group, which was injected with the nonspecific Silencer Negative Control #1 siRNA.
Figure 1. siRNA-induced Reduction of fLuc Protein and mRNA Levels in Mouse Liver. Ambion pMIR-REPORT™ Luciferase and β-galactosidase vectors and siRNA (1 or 5 nmol) that targets firefly luciferase (fLuc) were injected at constant high pressure into the tail veins of seven mice. Data were normalized to the average result from four mice injected with Silencer® Negative Control #1 siRNA. Panel A: fLuc and β-galactosidase (β-gal) protein levels were measured using the Tropix® Dual-Light® Luminescent Reporter Gene Assay System (Applied Biosystems). Each bar represents one animal, and readings were performed in triplicate. Panel B: TaqMan® Gene Expression Assays were used to measure fLuc mRNA levels. Each bar represents one animal, and readings were performed in triplicate and averaged.
The data demonstrated the following:
- The pMIR-REPORT Luciferase miRNA Expression Reporter Vector (as is, or with cloned targets) can be used in vivo and delivered by hydrodynamic injection to the mouse liver.
- siRNA can cause specific and potent knockdown of gene expression from a co-delivered reporter vector in liver cells.
- The Dual-Light Luminescent Reporter Gene Assay System can detect fLuc and β–gal protein in crude tissue samples.
Improving In Vivo Delivery of siRNAs
Co-delivery of siRNA with reporter vectors by hydrodynamic tail vein injection provides a starting point for researchers interested in performing RNAi experiments in the liver. There is a need to develop alternative techniques to enable specific delivery to other organs using normal low-pressure injection conditions.
Hydrodynamic tail vein injections were conducted under a license grant from Mirus Bio Corporation.
Scientific Contributors
Alexander V Vlassov, Mu Li, Susan Magdaleno • Applied Biosystems, Austin, TX
References
Hydrodynamic tail vein injections were conducted under a license grant from Mirus Bio Corporation.
Scientific Contributors
Alexander V Vlassov, Mu Li, Susan Magdaleno • Applied Biosystems, Austin, TX
References
- Tolentino MJ, Brucker AJ, Fosnot J, Ying GS, Wu IH, Malik G, Wan S, Reich SJ (2004) Intravitreal injection of vascular endothelial growth factor small interfering RNA inhibits growth and leakage in a nonhuman primate, laser-induced model of choroidal neovascularization. Retina 24:132–138.
- Tompkins SM, Lo CY, Tumpey TM, Epstein SL (2004) Protection against lethal influenza virus challenge by RNA interference in vivo. Proc Natl Acad Sci USA 101(23):8682–8686.
- Bitko V, Musiyenko A, Shulyayeva O, Barik S (2005) Inhibition of respiratory viruses by nasally administered siRNA. Nat Med 11(1):50–55.
- Zhang G, Budker V, Wolff JA (1999) High levels of foreign gene expression in hepatocytes after tail vein injections of naked plasmid DNA. Hum Gene Ther 10(10):1735–1737.
MethodsHydrodynamic (High-Pressure) Tail Vein Injections
The pMIR-REPORT™ Luciferase miRNA Expression Reporter Vector (5 µg) and pMIR-REPORT β-galactosidase Reporter Control Vector (5 µg) were mixed in 2.5 mL PBS and injected at constant high pressure into the tail veins of mice within 5–8 sec. For knockdown assays, Silencer® siRNA (~14 and 70 µg, equivalent to 1 and 5 nmol, respectively) targeting firefly luciferase (fLuc) was co-injected with the plasmids. The siRNA is expected to knock down fLuc expression, while expression of β-galactosidase (β-gal) from the control vector, used to monitor injection and nonspecific effects, should not change.
Analysis of Gene Expression and Knockdown
Liver, kidneys, spleen, and lung samples were collected from animals 24 hours after injection of plasmids and siRNA. Samples for protein analysis were homogenized in lysis solution from the Tropix® Dual-Light® Luminescent Reporter Gene Assay System (Applied Biosystems). Samples for RNA analysis were preserved in the Ambion RNA later® Tissue Collection: RNA Stabilization Solution at 4°C overnight, and total RNA was isolated with the Ambion mirVana™ PARIS™ Kit the next day.
Luciferase and β-galactosidase activity were measured using the Tropix Dual-Light Luminescent Reporter Gene Assay System. While this chemiluminescent reporter gene system is commonly used for assays in extracts from cultured cells, in this study it was successfully used to measure activity of fLuc and β-gal in crude tissue lysates, which had been neither purified nor enriched.
Levels of fLuc mRNA were measured by real-time RT-PCR using the TaqMan® Gene Expression Assay for fLuc. The RNA samples were reverse transcribed (Applied Biosystems High Capacity cDNA Reverse Transcription Kit), and the resulting cDNA samples were assayed with the TaqMan Universal PCR Master Mix, TaqMan primers and probe for fLuc, and the 7900HT Fast Real-Time PCR System. The relative level of gene expression for each sample was calculated by the ΔΔC T method.
The pMIR-REPORT™ Luciferase miRNA Expression Reporter Vector (5 µg) and pMIR-REPORT β-galactosidase Reporter Control Vector (5 µg) were mixed in 2.5 mL PBS and injected at constant high pressure into the tail veins of mice within 5–8 sec. For knockdown assays, Silencer® siRNA (~14 and 70 µg, equivalent to 1 and 5 nmol, respectively) targeting firefly luciferase (fLuc) was co-injected with the plasmids. The siRNA is expected to knock down fLuc expression, while expression of β-galactosidase (β-gal) from the control vector, used to monitor injection and nonspecific effects, should not change.
Analysis of Gene Expression and Knockdown
Liver, kidneys, spleen, and lung samples were collected from animals 24 hours after injection of plasmids and siRNA. Samples for protein analysis were homogenized in lysis solution from the Tropix® Dual-Light® Luminescent Reporter Gene Assay System (Applied Biosystems). Samples for RNA analysis were preserved in the Ambion RNA later® Tissue Collection: RNA Stabilization Solution at 4°C overnight, and total RNA was isolated with the Ambion mirVana™ PARIS™ Kit the next day.
Luciferase and β-galactosidase activity were measured using the Tropix Dual-Light Luminescent Reporter Gene Assay System. While this chemiluminescent reporter gene system is commonly used for assays in extracts from cultured cells, in this study it was successfully used to measure activity of fLuc and β-gal in crude tissue lysates, which had been neither purified nor enriched.
Levels of fLuc mRNA were measured by real-time RT-PCR using the TaqMan® Gene Expression Assay for fLuc. The RNA samples were reverse transcribed (Applied Biosystems High Capacity cDNA Reverse Transcription Kit), and the resulting cDNA samples were assayed with the TaqMan Universal PCR Master Mix, TaqMan primers and probe for fLuc, and the 7900HT Fast Real-Time PCR System. The relative level of gene expression for each sample was calculated by the ΔΔC T method.
Ambion Silencer® In Vivo Ready siRNAIn Vivo Ready
Silencer Pre-designed siRNA is subjected to the rigorous purification and testing required for introduction of siRNA into animals. After HPLC purification and annealing, each siRNA is dialyzed to remove salt (to <1 microsiemens conductance at 50 µM siRNA), sterile filtered, and endotoxin tested. This purification enables dosing strategies in mice of up to 50 mg/kg. In Vivo Ready
Silencer siRNA is the highest quality siRNA available anywhere.
Applications of the Ambion pMIR-REPORT™ miRNA Expression Reporter Vector SystemThis validated miRNA expression reporter vector system contains two mammalian expression vectors:
- pMIR-REPORT Luciferase miRNA Expression Reporter Vector contains firefly luciferase (fLuc) under control of a mammalian promoter/terminator system and a target cloning region downstream of the luciferase translation sequence. This vector was designed for cloning and testing of putative miRNA binding sites, for evaluation of endogenous miRNA expression, and for monitoring the effects of miRNA mimics (i.e., Ambion Pre-miR™ miRNA Precursors) or miRNA inhibitors (i.e., Ambion Anti-miR™ miRNA Inhibitors). In addition, a gene or viral target of interest can be cloned into this vector for in vivo and in vitro studies.
- pMIR-REPORT β-Galactosidase Reporter Control Vector is used for normalization of transfection efficiency.
Tropix® Dual-Light® Luminescent Reporter Gene Assay System for Luciferase and β-GalactosidaseThe
Dual-Light System can be used to quantitate firefly luciferase (fLuc) and β-galactosidase (β-gal) activity in the same sample. First, luciferase reporter enzyme activity is quantitated with an enhanced luciferase reaction. Following a 30–60 minute incubation and addition of a light emission accelerator, β-galactosidase reporter enzyme activity is quantitated with Galacton-Plus® substrate. The wide dynamic range of this dual assay enables accurate measurement of fLuc and β-gal concentrations over seven orders of magnitude (femtogram to nanogram range).