Search
Imaging a Dynamic Cellular Scaffold
In addition to acting as a cellular scaffold, the cytoskeleton has a role in organelle transport, cell division, motility, and signaling, making it central in both normal and disease processes. To facilitate research on the cytoskeleton, we offer phalloidin conjugates, CellLight and TubulinTracker reagents, and antibodies that react with the three main cytoskeletal subclasses—microfilaments, intermediate filaments, and microtubules—to enable fluorescence imaging of both live and fixed cells.
Actin and Microfilaments
Microfilaments, made up of linear actin polymers called F-actin, are the thinnest (6 nm diameter) and most dynamic of the three cytoskeletal subclasses. The polymers generate force by elongation at one end and shrinkage at the other. The actin monomer (G-actin) is a highly conserved protein used to form both microfilaments and the thin filaments of muscle cells. Actin participates in many important cellular processes, such as cell motility, phagocytosis, cytokinesis and division, organelle movement, muscle contraction, cell signaling, and establishment and maintenance of cell junctions and cell shape. Many of these processes are mediated by interactions of actin with cell membranes. In non-muscle cells, movement of cargo, such as organelles, along actin filaments is mediated by myosin-based motors in an ATP-dependent process.
Phallotoxins, such as phalloidin and phallacidin derivatives, bind competitively to F-actin. Fluorescent and biotinylated phallotoxins allow labeling of F-actin in fixed cells and can be used interchangeably in most applications. They stain F-actin at nanomolar concentrations and are water soluble, providing convenient probes for labeling, identifying, and quantitating F‑actin. Unlike antibodies, the binding affinity of phalloidin conjugates for actin does not change appreciably with actin from different species or sources; binding is in a stoichiometric ratio of about one phallotoxin molecule per actin subunit. Nonspecific staining is negligible, with great contrast between stained and unstained areas.
Because the phallotoxin conjugates are small (molecular weight <2,000), many actin-binding proteins—including myosin, tropomyosin, troponin, and DNase I—still bind to actin after treatment with phallotoxins. More significantly, phallotoxin-labeled actin filaments remain functional; labeled glycerinated muscle fibers still contract, and labeled actin filaments still move on solid-phase myosin substrates. Unlabeled phallotoxins may be used as controls to block F‑actin staining or promote actin polymerization (Figure 1).
While the Alexa Fluor phalloidin conjugates are unsurpassed for imaging fixed cells, they are membrane impermeant and thus unsuitable for live-cell analyses unless microinjected. For live-cell applications, BacMam technology can be used to introduce CellLight reagents—fluorescent proteins encoded by a nonreplicating baculovirus—for efficient expression of these proteins in living cells, including stem cells, neurons, and primary cells. Fluorescent actin and talin conjugates are both useful in studies of microfilaments. Talin is a cytoskeletal protein primarily found in regions of cell–substratum contacts, and can link integrins to the actin cytoskeleton directly or through interactions with vinculin and α-actinin. CellLight Actin GFP and CellLight® Talin GFP/RFP, like our other BacMam reagents, can be titered to optimal expression levels. Fluorescence is preserved after formaldehyde fixation, enabling staining with phalloidin conjugates or antibodies in multiplex fixed-cell analysis.
Due to the many cellular functions that involve microfilaments, research in the field spans a wide range of applications and topics. Examples of the use of Alexa Fluor phalloidin and secondary antibody conjugates include identification of the G-protein subunit responsible for integrin-mediated “outside-in” signaling [1] and studies of macular and retinal degeneration [2,3], and the role of the c-Jun N-terminal kinase (JNK1) in lentiviral entry in mammalian cells [4]. Cancer-related applications include modeling breast cancer tumor progression [5], demonstrating the role of actin dynamics in metastasis of epithelial cancer cells [6], and identifying a role for Lin28 in germ-cell development and malignancy [7]. Other applications include engineering of neuronal growth cones to promote axon regeneration [8] and studies of the influence of mechanical stimuli on embryonic stem cell differentiation [9].

Figure 1. Visualization of F-actin in fixed, permeabilized bovine pulmonary artery endothelial cells using Alexa Fluor phalloidin conjugates. Nuclei and F-actin were stained, respectively, with (A) DAPI and Alexa Fluor 680 phalloidin, (B) SYTOX Green dye and Alexa Fluor 568 phalloidin, (C) 7-AAD and Alexa Fluor 488 phalloidin, and (D) TO-PRO-3 iodide and Alexa Fluor 350 phalloidin. DAPI, SYTOX Green dye, 7-AAD, and TO-PRO-3 iodide are components of the SelectFX Nuclear Labeling Kit.
Intermediate Filaments
Intermediate filaments, about 10 nm in diameter, are a heterogeneous class of proteins that impart structure and function to the cytoskeleton. Much like actin, intermediate filaments maintain cell shape by bearing tension and imparting rigidity; intermediate filaments organize the internal three-dimensional structure of the cell, anchor organelles and the nucleus in place, and help form some cell–cell and cell–matrix junctions. Because intermediate filaments are abundant in cells subjected to mechanical stress, their primary role appears to be strengthening cells and tissues. Intermediate filaments are also structural components of the nuclear lamina that lines the inside of the nuclear membrane and governs the shape of the nucleus. Nuclear lamins help attach the chromosomes to the nuclear membrane and anchor nuclear pores.
Intermediate filament presence and composition is species dependent and varies with tissue type. Most mammalian epithelial cells contain keratins, while mesenchymal and muscle cells are rich in the fibrous proteins vimentin and desmin, respectively. Intermediate filaments of neurons and glial cells include peripherin, neurofilaments, and glial fibrillary acidic protein. A range of associated proteins bind to intermediate filaments, improving stability or providing attachment sites for other proteins such as actin filaments and microtubules.
Due to their structural and functional diversity, intermediate filaments are primarily imaged by antibody-based methods. We offer a large number of primary antibodies that recognize intermediate filaments in neurons, and other antibodies to cellular structures. Alexa Fluor dye–conjugated secondary antibodies offer many advantages for fluorescence imaging applications. They are very bright, are resistant to photobleaching and pH changes, and have narrow emission peaks for minimal bleedthrough. The quality of Alexa Fluor secondary antibodies has been demonstrated in thousands of high-impact publications.
Tubulin and Microtubules
Tubulins are a family of proteins, with α-tubulin and β-tubulin the most important members. Microtubules, assembled from dimers of α- and β-tubulin, are the largest of the cytoskeletal polymers, with a diameter of about 23 nm. Microtubules form from tubulin dimers by GTP-dependent polymerization. Dimers bound to GTP tend to assemble into microtubules, while dimers bound to GDP tend to fall apart; thus, this GTP cycle is essential for the dynamic instability of microtubules.
Microtubules play a key role in mitosis and chromosomal spindle formation, with anchoring points in the centrosome. Other important roles of microtubules in mammalian cells include intracellular movement of organelles, like mitochondria, in association with dyneins and kinesins. Other structures composed of microtubules are cellular flagella and cilia. Because of their role in mitotic cell division, tubulins are targets for anticancer drugs (taxol or paclitaxel, nocodazole, vinblastine, and vincristine) and certain anti-inflammatory agents (colchicine), which bind tubulin and affect its polymerization.
Tubulin Tracker or CellLight reagents can capture cytoskeletal dynamics and processes in live cells. Tubulin Tracker Green reagent, a fluorescent derivative of taxol, allows green-fluorescent staining of polymerized tubulin in live cells [10,11]. Tubulin Tracker Green reagent fluoresces when it enters live cells; because taxol binds polymerized tubulin, TubulinTracker Green reagent will inhibit cell division and possibly other functions utilizing polymerized tubulin in live cells.
Complementing Tubulin Tracker Green are CellLight tubulin and CellLight MAP4 reagents. Microtubule-associated protein 4 (MAP4), a major non-neuronal microtubule-associated protein, is an alternative to fluorescent protein conjugates of tubulin. Fusion of tubulin and MAP4 to fluorescent proteins like GFP and RFP, as in CellLight reagents, does not affect their in vivo function. CellLight actin and CellLight MAP4 reagents are delivered to cells using BacMam technology, so co-transducing cells with more than one CellLight reagent is easy (Figures 2 and 3). Typically, transduced cells express fusion proteins for about 5 days, though in slowly dividing cells, expression may last up to 2 weeks. In terminally differentiated neurons, images have been recorded more than 3 weeks after transduction.

Figure 2. Tracking of a cell undergoing mitosis with CellLight reagents. U2OS cells were transduced with CellLight MAP4-GFP (green) and CellLight Histone 2B–RFP (red) and imaged over 24 hours.
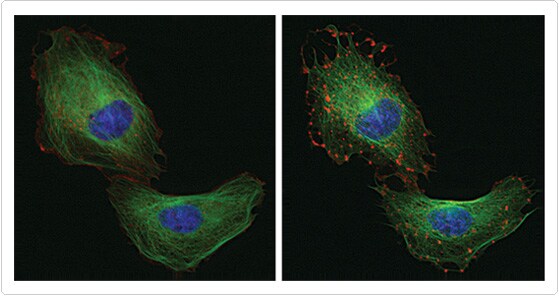
Figure 3. Multiplex imaging of microtubules and microfilaments. U2OS cells expressing CellLight MAP4-GFP and Cellular Lights Actin-RFP were labeled with 1 µg/mL Hoechst 33342 to stain nuclei. Cells were treated with 10 µM cytochalasin D and imaged at start of treatment and after 30 minutes. Cytochalasin D selectively disrupts actin filaments (red) without affecting microtubules (green).
References
- Goneg H, Shen B, Flevaris P et al. (2010) Science 327:340–343.
- Takeda A, Baffi JZ, Kleinman ME et al. (2009) Nature 460:225–230.
- Osakada F, Jin ZB, Hirami Y et al. (2009) J Cell Sci 122:3169–3179.
- Lee MH, Padmashali R, Andreadis ST (2010) J Virol 85:2657–2665.
- Han HJ, Russo J, Kohwi Y et al. (2008) Nature 452:187–193.
- Shankar J, Messenberg A, Chan J et al. (2010) Cancer Res 70:3780–3790.
- West JA, Viswanathan SR, Yabuuchi A et al. (2009) Nature 460:909–913.
- Hur EM, Yang IH, Kim DH et al. Proc Natl Acad Sci U S A 108:5057–5062.
- Hemsley AL, Hernandez D, Mason C et al. (2011) Cell Health Cytoskel 3:23–34.
- An S, Deng Y, Tomsho JW et al. (2010) Proc Natl Acad Sci U S A 107:12872–12876..
- Dikovskaya D, Li Z, Newton IP et al. (2010) J Cell Sci 123:736–746..
More Information
- Learn More About Phalloidin Conjugates
- Learn More About CellLight Fluorescent Protein Conjugates
- Check Out Our Cell Staining Tool
- Find Primary and Secondary Antibodies

Download This Article
Get a copy of this article as it appears in the print version of BioProbes 65.
Quick Product View
See a complete listing of the products discussed in this article.
View Products
For Research Use Only. Not for use in diagnostic procedures.