Search
Detecting miRNA & siRNA
Sensitive Solution Hybridization Assay For Small RNAs
In the past few years, interest in the identification, detection, and use of small RNA molecules has exploded. This interest has primarily stemmed from two interrelated lines of research. In one, small double-stranded RNAs (dsRNAs) called small interfering RNAs (siRNAs) have been used to silence the expression of specific genes at the post transcriptional level by a pathway known as RNA interference (RNAi). In the other, numerous small regulatory RNA molecules, referred to as microRNAs (miRNAs), have been shown to regulate target gene expression in various organisms. It is now becoming apparent that siRNAs and miRNAs are related molecules, sharing common processing pathways and maybe even functional mechanisms.
Currently, most miRNA researchers are analyzing miRNA expression patterns by Northern blot, a technique that is relatively insensitive and labor intensive. A few researchers performing gene silencing experiments also use this technique to analyze siRNA levels after RNAi induction, although the majority of researchers performing gene silencing experiments do not monitor siRNA levels at all. This may be due in large part to the inherent difficulties in detecting small RNAs with standard techniques. Here we discuss an optimized technique for the detection and quantitation of small RNAs. This technique allows easier monitoring of miRNA expression and siRNA levels in a variety of sample types and is considerably more sensitive than Northern analysis.
In the past few years, interest in the identification, detection, and use of small RNA molecules has exploded. This interest has primarily stemmed from two interrelated lines of research. In one, small double-stranded RNAs (dsRNAs) called small interfering RNAs (siRNAs) have been used to silence the expression of specific genes at the post transcriptional level by a pathway known as RNA interference (RNAi). In the other, numerous small regulatory RNA molecules, referred to as microRNAs (miRNAs), have been shown to regulate target gene expression in various organisms. It is now becoming apparent that siRNAs and miRNAs are related molecules, sharing common processing pathways and maybe even functional mechanisms.
Currently, most miRNA researchers are analyzing miRNA expression patterns by Northern blot, a technique that is relatively insensitive and labor intensive. A few researchers performing gene silencing experiments also use this technique to analyze siRNA levels after RNAi induction, although the majority of researchers performing gene silencing experiments do not monitor siRNA levels at all. This may be due in large part to the inherent difficulties in detecting small RNAs with standard techniques. Here we discuss an optimized technique for the detection and quantitation of small RNAs. This technique allows easier monitoring of miRNA expression and siRNA levels in a variety of sample types and is considerably more sensitive than Northern analysis.
Solution Hybridization Improves Sensitivity
Because hybridization of probe to target in solution is more sensitive than hybridization on solid support (Northern analysis), Ambion investigated the solution based RNase protection assay for its ability to detect small RNAs. After modification of the hybridization conditions to maximize hybridization of short probes to their targets, the assay's ability to specifically detect small RNA molecules were analyzed.
To examine the sensitivity of the assay, decreasing amounts of a 21 nt RNA, corresponding to the antisense strand of an siRNA known to target GAPDH, were spiked into a yeast RNA sample and then analyzed by the solution hybridization method. In brief, the samples were mixed with a 29 nt high specific activity radiolabeled probe (5 x 104 cpm) and hybridization buffer. After heat denaturation, each mixture was incubated at 42°C to hybridize the probe to the complementary siRNA target strand. Unhybridized RNA species and excess RNA probe were removed by a brief ribonuclease digestion. Protected target RNA fragments were recovered in the same tube using a reagent that simultaneously inactivates the ribonuclease and precipitates the nucleic acid. The protected RNA was then resuspended with gel loading buffer, analyzed on a denaturing polyacrylamide gel, and exposed to film. Figure 1A demonstrates that the assay was able to detect as little as 50 attomoles of the 21 nt target RNA after a two hour exposure.
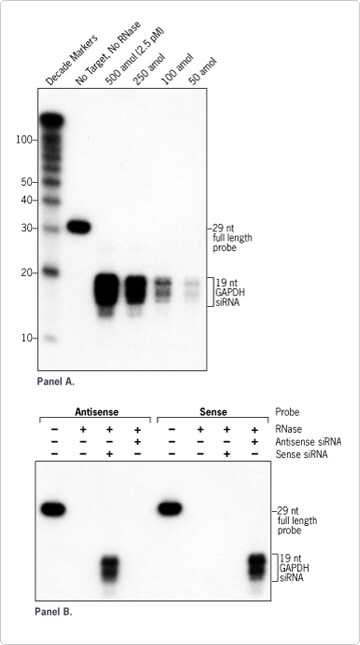
Figure 1. Sensitive and Specific Detection of MicroRNAs. (A) The indicated amounts of a 21 nt antisense GAPDH siRNA were spiked into 4 µg of yeast RNA and detected with the mirVana™ miRNA Detection Kit using a 29 nt long probe prepared with the mirVana miRNA Construction Kit. Protected RNA fragments (19 nt) were analyzed on a 15% denaturing polyacrylamide gel. (B) The same experiment as in Panel A with 200 attomoles of sense or antisense GAPDH siRNA radiolabeled probes specific for each strand..
To examine the sensitivity of the assay, decreasing amounts of a 21 nt RNA, corresponding to the antisense strand of an siRNA known to target GAPDH, were spiked into a yeast RNA sample and then analyzed by the solution hybridization method. In brief, the samples were mixed with a 29 nt high specific activity radiolabeled probe (5 x 104 cpm) and hybridization buffer. After heat denaturation, each mixture was incubated at 42°C to hybridize the probe to the complementary siRNA target strand. Unhybridized RNA species and excess RNA probe were removed by a brief ribonuclease digestion. Protected target RNA fragments were recovered in the same tube using a reagent that simultaneously inactivates the ribonuclease and precipitates the nucleic acid. The protected RNA was then resuspended with gel loading buffer, analyzed on a denaturing polyacrylamide gel, and exposed to film. Figure 1A demonstrates that the assay was able to detect as little as 50 attomoles of the 21 nt target RNA after a two hour exposure.
Figure 1. Sensitive and Specific Detection of MicroRNAs. (A) The indicated amounts of a 21 nt antisense GAPDH siRNA were spiked into 4 µg of yeast RNA and detected with the mirVana™ miRNA Detection Kit using a 29 nt long probe prepared with the mirVana miRNA Construction Kit. Protected RNA fragments (19 nt) were analyzed on a 15% denaturing polyacrylamide gel. (B) The same experiment as in Panel A with 200 attomoles of sense or antisense GAPDH siRNA radiolabeled probes specific for each strand..
Specificity of the Assay
Any assay for small RNAs must show specificity for the target molecule. Thus, the assay's specificity was investigated. As shown in Figure 1B, radiolabeled probes specific for the antisense strand of GAPDH siRNA detected only the corresponding target RNA. No signal was detected in the absence of target or in the presence of the sense strand of the GAPDH siRNA. Furthermore, the assay yielded no detectable background (e.g. nonspecific hybridization to yeast RNA molecules).
To further investigate assay specificity, miR-16 miRNA was detected in mouse kidney total RNA using three different probes. One probe was perfectly complementary to the target (miR-16), a second included three mismatched nucleotides (mir-16 mut), and a third included four additional A residues between the 22 nt sequence complementary to mir-16 and the leader sequence (mir-16 + 4). The mir-16 and mir-16 + 4 probes were both able to detect miR-16 miRNA in the RNA sample, whereas the signal was completely abolished with mir-16 mut probe (Figure 2). This result demonstrates that the assay has the required specificity for detecting small RNAs in total RNA samples.
To further investigate assay specificity, miR-16 miRNA was detected in mouse kidney total RNA using three different probes. One probe was perfectly complementary to the target (miR-16), a second included three mismatched nucleotides (mir-16 mut), and a third included four additional A residues between the 22 nt sequence complementary to mir-16 and the leader sequence (mir-16 + 4). The mir-16 and mir-16 + 4 probes were both able to detect miR-16 miRNA in the RNA sample, whereas the signal was completely abolished with mir-16 mut probe (Figure 2). This result demonstrates that the assay has the required specificity for detecting small RNAs in total RNA samples.
Detecting Multiple Small RNAs in the Same Sample
Figure 2 also shows that the assay can be used for multi-target detection. The four A residues added to the mir-16 probe between the complementary region and the leader sequence (which is necessary to distinguish protected from unprotected probe) are not cleaved by the mixture of RNases used in the assay. They thus increase the size of the protected fragment by four nucleotides. This probe (mir-16 +4) can be used in the presence of another probe with the same length complementary region but lacking the additional A residues (mir-22) for multi-target detection (Figures 2 and 3). Adding multiple A residues to a small RNA probe sequence permits the detection of more than one small RNA in a single sample giving this assay a distinct advantage over Northern analysis for detecting multiple small RNA molecules that have the same size.
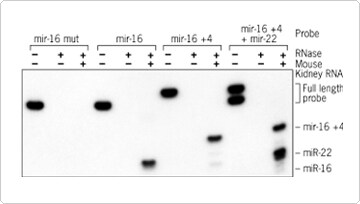
Figure 2. miRNA Expression in Mouse Kidney Total RNA. miR-16 and miR-22 expression was analyzed as in Figure 1 with 1 µg of FirstChoice Total RNA from mouse kidney and 32 nt long probes generated with the mirVana miRNA Probe Construction Kit. mir-16 mut probe (32 nt) carries 3 mismatch mutations (ACG to CGA) corresponding to nucleotides 9 to 11 of the miR-16 miRNA sequence. The mir-16+4 probe (36 nt) carries 4 additional A residues between the 22 nt sequence specific for miR-16 and the 10 nt leader sequence, producing a 26 nt long protected fragment, which is 4 nt longer than that produced by the mir-16 and mir-22 probes.
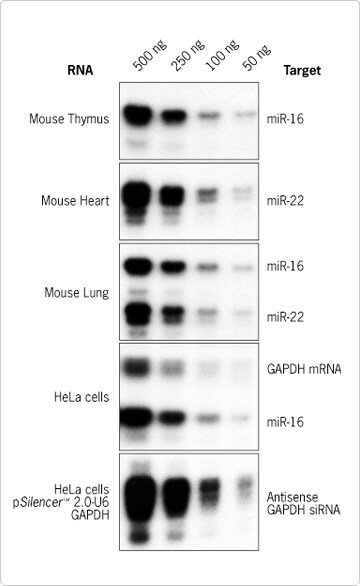
Figure 3. Single and Multiple Target Detection. The indicated target RNAs were detected in 500, 250, 100 and 50 ng of total RNA from mouse tissues (3.5 hours exposure) or HeLa cells (6 hours exposure) as described in Figures 1 and 2. The probe specific for GAPDH mRNA (39 nt) produces a 29 nt long protected fragment with the same specific activity as the mir-16 protected fragment. miR-16 was detected with the mir-16 +4 probe.
Figure 2. miRNA Expression in Mouse Kidney Total RNA. miR-16 and miR-22 expression was analyzed as in Figure 1 with 1 µg of FirstChoice Total RNA from mouse kidney and 32 nt long probes generated with the mirVana miRNA Probe Construction Kit. mir-16 mut probe (32 nt) carries 3 mismatch mutations (ACG to CGA) corresponding to nucleotides 9 to 11 of the miR-16 miRNA sequence. The mir-16+4 probe (36 nt) carries 4 additional A residues between the 22 nt sequence specific for miR-16 and the 10 nt leader sequence, producing a 26 nt long protected fragment, which is 4 nt longer than that produced by the mir-16 and mir-22 probes.
Figure 3. Single and Multiple Target Detection. The indicated target RNAs were detected in 500, 250, 100 and 50 ng of total RNA from mouse tissues (3.5 hours exposure) or HeLa cells (6 hours exposure) as described in Figures 1 and 2. The probe specific for GAPDH mRNA (39 nt) produces a 29 nt long protected fragment with the same specific activity as the mir-16 protected fragment. miR-16 was detected with the mir-16 +4 probe.
Analysis of miRNA Expression Patterns Across Various Tissue Types
The function of most miRNAs is not known; however, a number of miRNAs seem to be involved in post-transcriptional gene regulation. Some of these miRNAs (e.g. lin-4 and let-7) inhibit protein synthesis by binding to the 3' untranslated region of target mRNAs. Others bind to perfectly complementary mRNA sequences to destroy target transcripts (e.g.
Scarecrow miRNA in plants). These miRNAs, therefore, function like siRNAs and could be classified as such.
Bantam,
lin-4 and
let-7 have also been shown to play critical roles in tissue development. Other miRNAs are believed to have similar functions because of their differential spatial and temporal expression patterns.
We used the solution hybridization assay to monitor the differential expression of two different miRNAs across mouse tissues (Figure 4). High levels of mir-16 expression were detected in all five tissues tested, with the highest expression levels evident in lung and thymus. These variations across tissues were confirmed by Northern blot analysis with the same mir-16 probe. Interestingly the mir-22 probe showed a completely different pattern of expression. miR-22 was highly expressed in lung and ovary and was present in spleen, thymus, and testicle at levels that would not have been detectable with standard Northern blotting techniques (data not shown). The relative abundance of miR-16 and miR-22 miRNA in mouse lung was also confirmed by multi-target detection with the simultaneous use of the mir-16+4 and mir-22 probes in the assay (Figure 3).
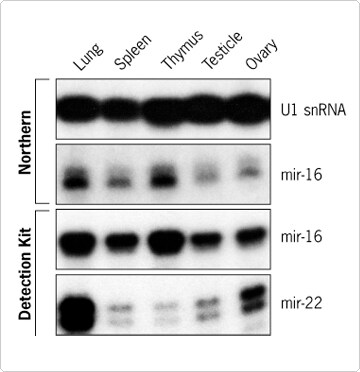
Figure 4. miR-16 and miR-22 Expression in Mouse Tissues. miR-16 and miR-22 miRNAs (22 nt) were detected in 1 µg of FirstChoice Total RNA from five different mouse tissues using 32 nt long mir-16 or mir-22 probes generated with the mirVana miRNA Probe Construction Kit. The same differential expression of miR-16 across tissues was observed by Northern blot analysis (2 days exposure) or by hybridization in solution (2 hr exposure). RNAs were analyzed on 15% denaturing polyacrylamide gels. As a loading control, the same RNA samples were resolved on a 1.2% denaturing agarose gel and U1 snRNA expression was analyzed by Northern blot.
We used the solution hybridization assay to monitor the differential expression of two different miRNAs across mouse tissues (Figure 4). High levels of mir-16 expression were detected in all five tissues tested, with the highest expression levels evident in lung and thymus. These variations across tissues were confirmed by Northern blot analysis with the same mir-16 probe. Interestingly the mir-22 probe showed a completely different pattern of expression. miR-22 was highly expressed in lung and ovary and was present in spleen, thymus, and testicle at levels that would not have been detectable with standard Northern blotting techniques (data not shown). The relative abundance of miR-16 and miR-22 miRNA in mouse lung was also confirmed by multi-target detection with the simultaneous use of the mir-16+4 and mir-22 probes in the assay (Figure 3).
Figure 4. miR-16 and miR-22 Expression in Mouse Tissues. miR-16 and miR-22 miRNAs (22 nt) were detected in 1 µg of FirstChoice Total RNA from five different mouse tissues using 32 nt long mir-16 or mir-22 probes generated with the mirVana miRNA Probe Construction Kit. The same differential expression of miR-16 across tissues was observed by Northern blot analysis (2 days exposure) or by hybridization in solution (2 hr exposure). RNAs were analyzed on 15% denaturing polyacrylamide gels. As a loading control, the same RNA samples were resolved on a 1.2% denaturing agarose gel and U1 snRNA expression was analyzed by Northern blot.
Simultaneous Detection of siRNA Expression and Target Gene Knockdown
For the development of improved siRNA delivery systems and siRNA expression vectors, particularly those that permit targeted delivery or tissue specific expression, researchers need to monitor siRNA levels -- preferably in conjunction with the assessment of target transcript down regulation. To determine if the solution based assay permitted correlation of expressed siRNA levels and targeted mRNA knockdown, probes were generated to a GAPDH siRNA known to effectively reduce GAPDH expression, and to the GAPDH mRNA target. An expression vector p
Silencer™ 2.0-U6) engineered to express either the GAPDH siRNA or a scrambled control siRNA was transfected into HeLa cells. Three days after transfection, total RNA was isolated and the solution hybridization assay was performed. Using the assay, it was possible to simultaneously detect siRNA expressed from the vector (Figures 3 and 5A), as well as the GAPDH mRNA (Figure 5B). As expected, the GAPDH mRNA levels were found to be reduced considerably (~60%) in cells transfected with the GAPDH siRNA vector as compared to cells transfected with the negative control vector (Figure 5B). In addition, the GAPDH siRNA was only detected in the cells transfected with the GAPDH siRNA expressing vector. The specific reduction of GAPDH mRNA expression was similar (~55%) when analyzed by Northern blot.
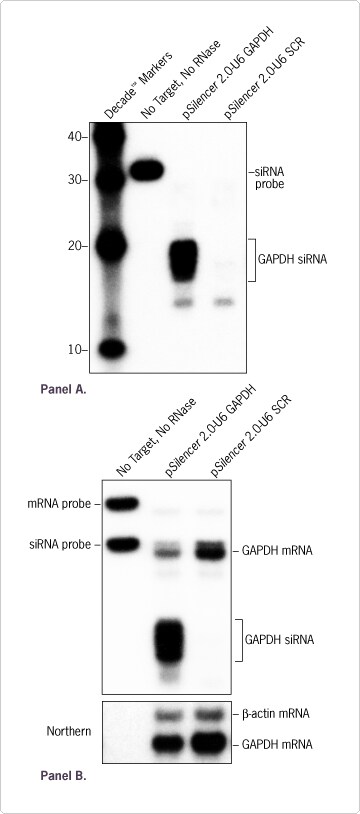
Figure 5. Analysis of GAPDH siRNA Expression and mRNA Knockdown. (A) HeLa cells were transfected with pSilencer 2.0-U6 engineered to express either an siRNA targeting GAPDH or a negative control siRNA (SCR). Three days after transfection, total RNA was isolated and 1 µg was assessed using the mirVana miRNA Detection Kit. Probes to the antisense strand of the GAPDH siRNA were prepared as described in Figure 1. (B) Same experiment with probes specific for GAPDH mRNA or GAPDH siRNA. Both probes had the same specific activity. As a control, GAPDH mRNA expression was also analyzed by Northern blot.
Figure 5. Analysis of GAPDH siRNA Expression and mRNA Knockdown. (A) HeLa cells were transfected with pSilencer 2.0-U6 engineered to express either an siRNA targeting GAPDH or a negative control siRNA (SCR). Three days after transfection, total RNA was isolated and 1 µg was assessed using the mirVana miRNA Detection Kit. Probes to the antisense strand of the GAPDH siRNA were prepared as described in Figure 1. (B) Same experiment with probes specific for GAPDH mRNA or GAPDH siRNA. Both probes had the same specific activity. As a control, GAPDH mRNA expression was also analyzed by Northern blot.
Advantages of the Solution Hybridization Assay
The experiments presented here indicate that the solution hybridization assay based on ribonuclease protection can sensitively and specifically detect small RNAs such as miRNAs and siRNAs. Quantitative analyses can be performed in solution with as little as 10-50 ng of total RNA to detect attomole (10-18 mol) amounts of target RNA. In general, solution hybridization with short RNA probes is 100 500 times more sensitive than membrane hybridization. Another advantage of this assay is the potential to simultaneously detect several small RNAs of the same size or both small RNA and longer RNA species (e.g. siRNA and target messenger RNA) from a single sample.
Ambion makes this solution hybridization assay available as the mirVana miRNA Detection Kit. We also offer the mirVana miRNA Probe Construction Kit, for the easy preparation of short RNA probes for this assay as well as for Northern analysis and in situ hybridization.
Ambion makes this solution hybridization assay available as the mirVana miRNA Detection Kit. We also offer the mirVana miRNA Probe Construction Kit, for the easy preparation of short RNA probes for this assay as well as for Northern analysis and in situ hybridization.
Decade™ Markers: The Ideal Markers for siRNA and miRNA AnalysisAmbion's Decade Marker System is ideal for research laboratories studying miRNA, siRNA, snRNA, and other small RNA molecules. The Decade Marker System produces a set of radiolabeled RNA molecules of 100, 90, 80, 70, 60, 50, 40, 30, 20, and 10 nucleotides (Figure 1). The system includes all of the reagents necessary to produce the molecular weight markers except [gamma 32P]ATP. The Decade Markers are generated from a single, gel-purified transcript that is provided. The transcript is 5' end-labeled using the T4 polynucleotide kinase and kinase buffer supplied with the kit. The phosphorylation reaction is then diluted into a cleavage reagent that generates the molecular weight marker in a five minute, room temperature reaction. Gel loading buffer from the kit is then added to the Decade Markers to prepare them for electrophoresis. The labeling, cleavage, and gel preparation steps can be completed in less than an hour. Each Decade Marker Kit contains reagents for 10 preparative reactions, providing RNA markers for as many as 200 experiments.
Solution Hybridization AssayAssays were performed with the mirVana miRNA Detection Kit as described in the Instruction Manual. RNA samples were mixed with the high specific activity radiolabeled probes (5 x 104 cpm) and the included 2X hybridization buffer. After heat denaturation, each mixture was incubated at 42°C to hybridize the probe to its complement. Unhybridized RNA and excess probe were then removed by a digestion using the included RNase mixture. Protected target RNA fragments were recovered in the same tube using the RNase Inactivation/Precipitation Reagent. Fragments were then resuspended in 5 µl gel loading buffer, separated on a 15% denaturing polyacrylamide gel, and detected by autoradiography. Only 5 10% of the "No target/No RNase" controls were loaded.
Northern AnalysisFor miRNA analysis, total RNA samples were electrophoresed on a 15% acrylamide gel, electrotransferred to positively charged nylon membrane (Ambion), and then hybridized at 42°C to the indicated probes using ULTRAhyb-Oligo hybridization solution. For analysis of GAPDH, ß-actin, and U1 mRNA, total RNA was separated on a 1.2% agarose gel and then detected with the indicated probes using the NorthernMax-Gly Kit (Ambion) according to the instruction manual.